Основы молекулярной биологии. Часть 2: Молекулярные генетические механизмы - А.Н. Огурцов 2011
Клонирование ДНК
Создание кДНК библиотек
Основные этапы создания кДНК библиотеки X фагов из смеси клеточных мРНК представлены на рисунке 91.
Для создания кДНК библиотеки необходимо сначала выделить интересующую мРНК из определённых клеток данной ткани.
мРНК достаточно просто отделить от присутствующих в клетках в намного больших количествах рРНК и тРНК, используя тот факт, что мРНК (в отличие от рРНК и тРНК) имеет поли-(А) хвост. Для отделения мРНК используют фильтр с присоединенными к нему олиго-(дТ) (короткие участки тимидилатов).
Для синтеза нитей ДНК, комплементарных мРНК, начиная с поли-(dТ) праймера (рисунок 91, этап 2), используют фермент обратная транскриптаза, который был найден у ретровирусов (п. 7.4.2).
Полученные гибридные молекулы мРНК-кДНК преобразовываются в несколько этапов в двухцепочечные молекулы кДНК, соответствующие всем мРНК молекулам, которые были в исходной смеси (этапы 3-5).
Каждая такая молекула кДНК имеет участок (олиго-dС)•(олиго-dG) на одном конце и (олиго-dА)-(олиго-dТ) на другом (рисунок 91, этап 5).
Для защиты кДНК от разрезания рестрикционными ферментами проводят метилирование кДНК (рисунок 91, этап 6).
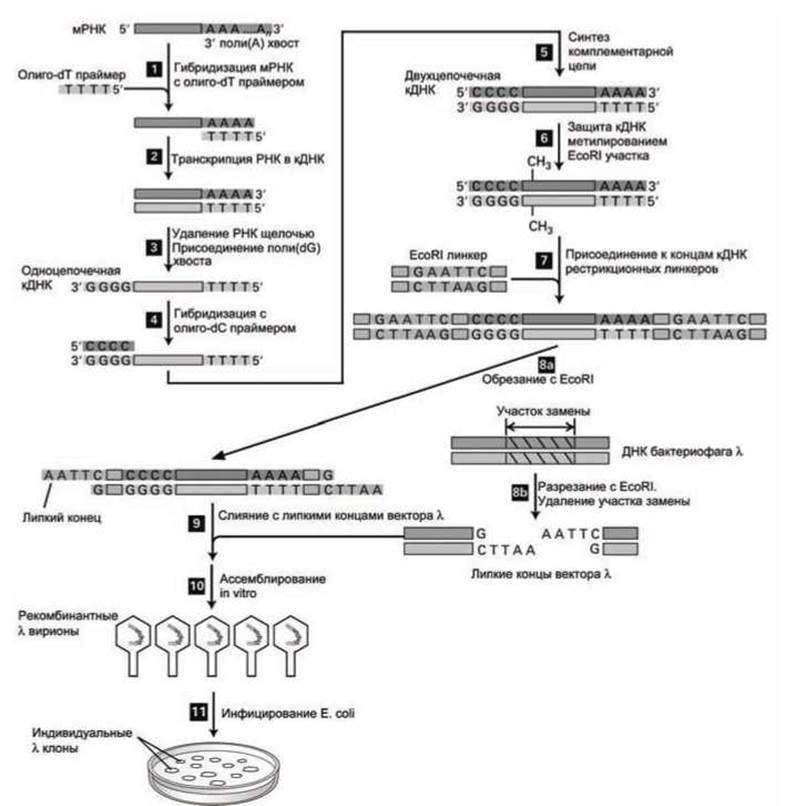
Рисунок 91 - Создание библиотеки кДНК с использованием бактериофага λ
Затем, подготавливая кДНК к клонированию, к обоим концам кДНК присоединяют с помощью ДНК-лигазы бактериофага Т4 короткие двухцепочечные молекулы ДНК, имеющие сайты рестрикции для определённого фермента рестрикции (рисунок 91, этап 7). На рисунке показан пример присоединения участков ДНК, содержащих сайт рестрикции для фермента ЕсоRI.
Как уже упоминалось ранее, ДНК-лигаза бактериофага Т4 может соединять "тупые" концы двухцепочечных ДНК, не имеющих "липких" концов.
Полученные таким образом ДНК-молекулы затем обрабатываются соответствующим рестрикционным ферментом (на рисунке, EcoRI). В результате получаются кДНК молекулы с липкими обоими концами (рисунок 91, этап 8а).
Одновременно с этим готовят "векторные руки" (vector arms) из геномов бактериофага λ, обрабатывая их тем же рестрикционным ферментом (в нашем примере EcoRI). Эти векторные руки имеют липкие концы и содержат в себе все гены, необходимые для литического роста вируса (рисунок 91, этап 8(6)). При этом, как правило, удаляют несущественный для создания библиотеки участок внедрения (рисунок 89, косая штриховка), который также называют участком замены (рисунок 91, участок с косой штриховкой на этапе 8(6)).
Набор кДНК и векторных рук, имеющих одинаковые липкие концы, смешивают и соединяют ковалентно с помощью ДНК-лигаз (рисунок 91, этап 9). Каждая из полученных рекомбинантных молекул ДНК содержит кДНК, расположенную между двумя векторными руками λ-вектора ДНК.
Затем, in vitro, ассемблируются (собираются, как показано на рисунке 90) вирионы, содержащие рекомбинантные ДНК (рисунок 91, этап 10). В состав вириона могут войти только молекулы, длина которых не превышает 49 kb.
И, наконец, рекомбинантные λ-фаги помещаются на посев Е. coli, чтобы вырастить индивидуальные бляшки (рисунок 91, этап 11).
Поскольку каждая бляшка происходит от единственного рекомбинантного фага, то все дочерние 1-фаги в этой бляшке генетически идентичны и представляют собой клон, содержащий кДНК, полученный из единственной мРНК. Весь набор клонов разных бляшек образует λ-кДНК библиотеку.
Есть одно свойство таких кДНК библиотек, которое связано с тем, что различные гены транскрибируются с различной скоростью (интенсивностью). В результате кДНК клоны, соответствующие быстро транскрибируемым генам, будут представлены в библиотеке многократно, в то время как медленно транскрибируемые гены будут встречаться редко или вообще будут отсутствовать.
Это свойство удобно использовать в тех случаях, когда исследуются именно гены, транскрибируемые с большой скоростью. Именно "обогащённость" библиотеки ДНК копиями таких генов облегчает скрининг такой библиотеки.
Однако, для того, чтобы обеспечить наличие в библиотеке также и клонов, соответствующих медленно транскрибируемым генам, библиотеки кДНК млекопитающих должны содержать 106—107 (до десяти миллионов) индивидуальных клонов рекомбинантных λ-фагов.
ВЫВОДЫ
Для клонирования ДНК создают in vitro рекомбинантные молекулы ДНК, помещая фрагменты ДНК в ДНК-векторы. Рекомбинантные молекулы ДНК затем помещают в клетку, где они реплицируются, образуя большое количество копий ДНК.
Рестрикционные ферменты (эндонуклеазы) обычно разрезают ДНК в специфических местах, длиной 4-8 пар оснований и имеющих па- линдромную структуру, образуя при этом определённые фрагменты ДНК, имеющие комплементарные одноцепочечные хвосты (липкие концы).
Два рестрикционных фрагмента с комплементарными концами могут быть объединены с помощью ДНК-лигазы.
Плазмидные векторы Е. coli представляют собой круговые молекулы ДНК, имеющие три необходимых функциональных участка: (1) ориджин репликации, (2) ген резистивности к антибиотику, и (3) участок, в котором фрагмент ДНК может быть включен в состав плазмиды. Из трансформированных клеток, имеющих в своем составе векторы, вырастают колонии в среде, которая обеспечивает селектирующий эффект.
Клонирующие векторы на основе фага Л образуются замещением несущественной части генома λ фрагментом ДНК длиной до 25 kb и ассемблированием полученной рекомбинантной ДНК с предварительно синтезированными in vitro головкой и хвостом фага.
При клонировании кДНК функциональная мРНК транскрибируется в комплементарную ДНК. После серии преобразований одноцепочечная кДНК конвертируется в двухцепочечную кДНК, которая затем встраивается в вектор фага λ.
Библиотека кДНК - это набор кДНК клонов, приготовленных из мРНК, полученных из определённого типа тканей. Геномная библиотека - это набор клонов, содержащих рестрикционные фрагменты, полученные при фрагментировании всего генома.
Число клонов в геномной или кДНК-библиотеке должно быть настолько большим, чтобы любая исходная нуклеотидная последовательность присутствовала хотя бы в одном из клонов.
Вопросы для самоконтроля
1. Что называется клонированием ДНК?
2. Какая ДНК называется рекомбинантной?
3. Что называется генетическим вектором?
4. Для чего в технологии рекомбинантных ДНК используют рестрикционные ферменты?
5. Для чего в технологии рекомбинантных ДНК используют ДНК-лигазы?
6. Какая последовательность нуклеотидов называется палиндромной?
7. Какой участок ДНК называется сайтом рестрикции?
8. Какова функция модифицирующего фермента в рестрикционно-модифицирующей системе бактерий?
9. Какие участки ДНК называются "липкими" концами? Как их получают и в чем их отличие от "тупых" концов?
10. Какие участки ДНК называются "тупыми" концами? Как их получают и в чем их отличие от "липких" концов?
11. Какие фрагменты ДНК называются рестрикционными фрагментами?
12. Что такое плазмиды?
13. Перечислите основные компоненты плазмидного клонирующего вектора.
14. Что такое полилинкер и для чего его используют при синтезе плазмидных векторов?
15. В чем заключается процесс генетической трансформации клеток, и какие клетки называются трансформированными?
16. В чем состоят преимущества использования бактериофага λ в качестве клонирующего вектора по сравнению с плазмидными клонирующими векторами?
17. Что называется конкатомером бактериофага λ и какова роль белков Nul и А в процессинге конкатомеров?
18. Что называется геномной библиотекой?
19. Какие ДНК называются комплементарными и как получают комплементарные ДНК?
20. В чем преимущество библиотеки комплементарных ДНК (кДНК библиотеки) по сравнению с простой геномной библиотекой?
21. Для чего используется фермент обратная транскриптаза при создании библиотеки комплементарных ДНК (библиотеки кДНК)?