Основы молекулярной биологии - В.И. Резяпкин 2009
Репликация
Механизм репликации у E.сoli
В клетках E.coli присутствуют три ДНК-полимеразы. Охарактеризуем их.
ДНК-полимераза I
На одной полипептидной цепи ДНК-полимеразы I находятся 2 активных центра:
а) 1-ый активный центр ответственен за полимеразную и 3’→5’-экзонуклеазную активности. Последняя обеспечивает удаление ошибочно встроенных нуклеотидов;
b) 2-ый активный центр ответственен за 5’→3’-экзонуклеазную активность. Эта активность необходима для удаления РНК-затравки в процессе репликации.
ДНК-полимераза II
Этот фермент обладает полимеразной и 3’→5’-экзонуклеазной активностями, предпочтительнее работает на двухцепочечных ДНК с брешами (рис. 2.9). ДНК-полимераза II участвует в репарации ДНК.
Рис. 2.9. ДНК-полимераза II предпочтительнее работает на двухцепочечных
ДНК с брешами
ДНК-полимераза III
Этот фермент обладает полимеразной и 3’→5’-экзонуклеазной активностями, состоит из десяти типов субъединиц. Его основное назначение - репликация ДНК. Скорость синтеза - 500 нуклеотидов в секунду
Хромосома E.coli имеет одну точку начала репликации (oriC), ее - размер 258 н.п. Белок Dna А узнает OriC и инициацирует репликацию. В результате АТФ-зависимая хеликаза начинает расплетать дуплекс ДНК (рис. 2.10). Топоизомераза, располагаясь впереди по ходу движения репликативной вилки, снимает напряжение, возникаюшее в результате расплетения двойной спирали в ДНК. С образующимися одноцепочечными участками ДНК связывается SSB-белок (рис. 2.10). Праймаза осуществляет синтез затравки (рис. 2.10, 2.11), которая необходима для проявления активности ДНК-полимеразы. Затем в работу включается ДНК-полимераза III, которая последовательно присоединяет нуклеотиды к 3’-концу полинуклетидной цепи (рис. 2.10). Поскольку синтез ДНК осуществляется в направлении 5’→3’, одна цепь (ведущая) синтезируется непрерывно (рис. 2.11), вторая фрагментами (отстающая цепь) по 1000 - 2000 нуклеотидов (фрагменты Оказаки). По окончанию синтеза фрагмента Оказаки ДНК-полимераза I за счет 5’→3’- экзонуклеазной активности удаляет затравку и заменяет ее ДНК. После действия этого фермента между фрагментами Оказаки остается разрыв, который сшивает ДНК-лигаза (рис.2.10, 2.11).
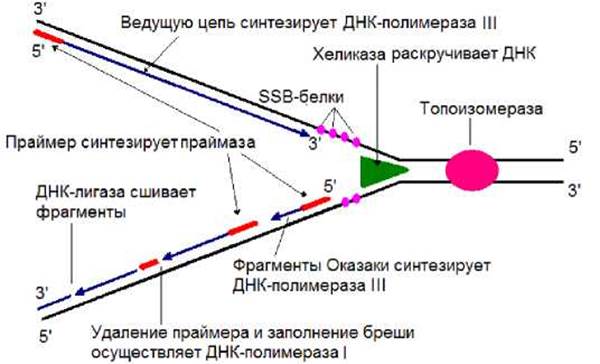
Рис. 2.10. Кооперативное действие белков в репликационной вилке
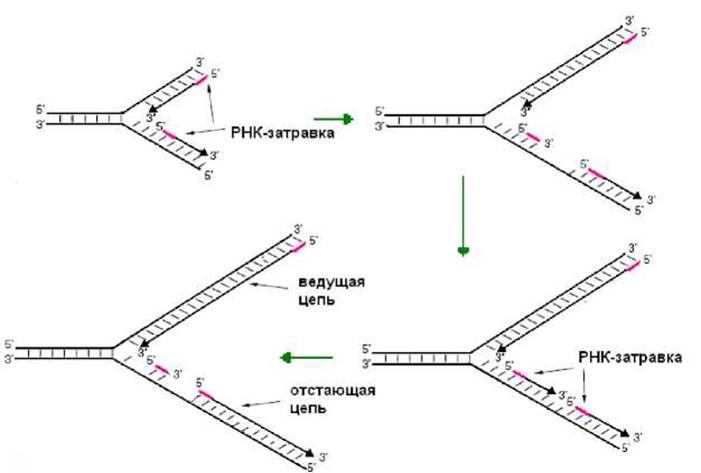
Рис. 11. Синтез цепей ДНК в процессе репликации
Терминация репликации происходит после удвоения кольцевой молекулы ДНК.