Основы молекулярной биологии - В.И. Резяпкин 2009
Репликация
Синтез теломер
Соматические клетки могут делиться ограниченное число раз, например, эмбриональные клетки могут делиться до 80 раз, клетки 70-летнего человека - только 20 - 30 раз. Способность клеток делится ограниченное число раз носит название лимита Хейфлика.
В ряде случаев клетки способны преодолеть лимит Хейфлика и начать делиться неограниченно, это явление называется - иммортализация клеток.
За 50 делений масса потомков одной клетки может составить 20 000 000 т.
Ограничение числа делений клеток определяется наличием особых структур на концах хромосом - теломер. Теломеры состоят из повторяющихся коротких последовательностей ДНК. У многих видов они представлены высококонсервативными повторами ТТАГГГ. ДНК теломер не кодирует белки. У человека размер теломер может составлять от 2000 до 20 000 п.н. В процессе развития организма размер теломер уменьшается. Клетки, имеющие меньший размер теломер, имеют меньший потенциал делений. Уменьшение теломер в процессе деления клеток связано с недорепликацией концов хромосом. Недорепликация концов хромосом определяется тем, что в процессе репликации используется РНК-затравка и синтез ДНК протекает в направлении 5’→3’ (рис. 2.14).
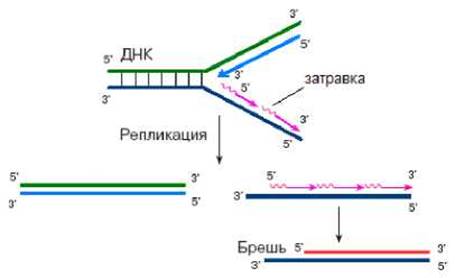
Рис. 2.14. Недорепликация хромосом
За наращивание теломер ответственен фермент - теломераза. Однако на определенной стадии развития происходит выключение гена теломеразы. Поэтому теломеры в процессе деления укорачиваются, что приводит к ухудшению функционирования хромосом, так как теломеры защищают генетический материал от деградации и ответственны за расположение хромосомы в клетке и ее функционирование. Следует отметить, что теломеразные гены выключаются только у организмов, размножающихся половым путем, но не вегетативным.
Теломераза является РНК-содержащим ферментом. В составе РНК-компонента теломеразы имеется тринуклеотид, комплементарный соответствующему участку повторяющейся последовательности теломеры. В результате их комплементарного взаимодействия (рис.2.15) образуется матричная область, которую использует фермент для синтеза фрагмента теломеры. После заверщения синтеза этого фрагмента происходит перемещение (транслокация РНК-компонента теломеразы в направлении 3’-конца, синтезируемой цепи. Теломераза вновь способна продолжить синтез ДНК. Этот циклический процесс повторяется многократно до тех пор, пока не завершится синтез соответствующей цепи ДНК.
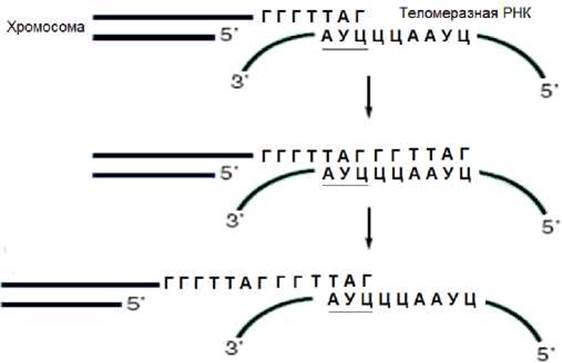
Рис. 2.15. Механизм действия теломеразы
После того как теломераза завершит синтез цепи ДНК в направление 3’-конца, при участии ДНК-полимеразы и других белков происходит синтез комплементарной ей цепи (рис. 2.15). Так происходит образование теломер.
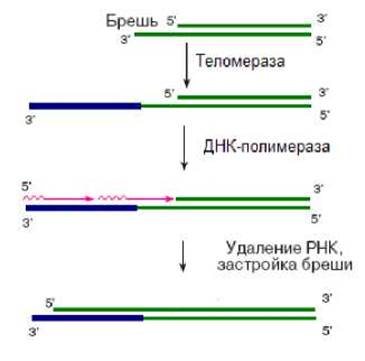
Рис. 2.15. Образование теломеры
Теломеразная РНК закодирована в генах, ее длина у различных организмов не одинакова: у простейших она составляет 150 - 200 нуклеотидов, у человека и мыши - 450 нуклеотидов, у дрожжей - 1300 нуклеотидов.
Матричная область РНК теломеразы находится на расстоянии 50 нуклеотидов от ее 5’-конца. У теломеразы мышей матричная область РНК составляет 8 нуклеотидов, у человека - 11. Тем не менее, обе последовательности кодируют одну и ту же повторяющуюся единицу теломеры.
В зародышевых клетках ген теломеразы экспрессируется, и, следовательно, в них проявляется теломеразная активность, и происходит синтез теломер. В соматических же клетках ген теломеразы выключен, и теломеразная активность не определяется.
В раковых клетках происходит активация гена теломеразы, и поэтому в них уровень активности теломеразы достаточно высок и вследствие этого происходит синтез теломер. Они в раковых клетках хоть и короче, чем в эмбриональных, но стабильны.
Использование антисмысловых РНК к РНК-компоненту теломеразы вызывало гибель опухолевых клеток HeLa. Эти эксперименты говорят о важной роли теломеразы в перерождении нормальных клеток в раковые. В тоже время получены мыши, нокаутированные по гену, кодирующему РНК-компонент теломеразы. У них отсутствовала активность теломеразы. Однако они были жизнеспособными в течение 6 поколений. Кроме того клеточные линии из этих мышей подвергались иммортализации, трансформировались вирусными онкогенами и становились онкогенными для бестимусных мышей. Клетки мышей четвертого поколения не содержали теломерных повторов, характеризовались различными аномалиями. Эти данные подтверждают необходимость теломеразы для поддержания теломер, но отрицают ее роль в иммортализации клеток и их злокачественной трансформации.
Подытоживая вышесказанное, следует отметить, что воспроизведение генетической информации является сложнейшим биологическим процессом, в котором участвуют многочисленные факторы.