Основы молекулярной биологии - В.И. Резяпкин 2009
Транскрипция
Транскрипция прокариот
Прокариоты содержат одну РНК-полимеразу, состоящую из нескольких субъединиц. Наиболее изучена РНК-полимераза E. coli. Этот фермент состоит из субъединиц, обозначаемых буквами греческого алфавита а, ß, ß’ и σ. РНК-полимераза может существовать в двух формах - в форме холофермента, его субъединичный состав выражается формулой a2ßß’σ, и корфермента - a2ßß’. Только холофермент может инициировать синтез РНК. После инициации транскрипции σ-субъединица отделяется и элонгацию осуществляет корфермент. Таким образом, σ-субъединица, или ее еще называют σ-фактор, необходима для узнавания промотора. ß-субъединица участвует в связывании НТФ, ß’-субъединица взаимодействует с ДНК, комплекс a2ß’ специфически связывается с промоторными нуклеотидными последовательностями. При взаимодействии с ДНК РНК-полимераза «закрывает» участок размером 60 п.н.
В клетке E.coli присутствуют несколько G-факторов, они ответственны за узнавание РНК-полимеразой различных промоторов. Как правило σ-факторы узнают блоки, отстоящие от точки начала транскрипции приблизительно на 10 и 35 нуклеотидов (рис. 4.3). Эти блоки имеют консервативные (общие для всех в пределах одной группы промоторов) нуклеотидные последовательности.
![]()
Рис.4.3. Организация промотора. Номером +1 обозначается нуклеотид матричной цепи, с которого начинается транскрипция. Нуклеотиды, стоящие перед ним, имеют отрицательные номера, после него - положительные. Последовательность ТАТААТ называется ТАТА-блоком или Прибнов-блоком. Последовательность ТТГАЦА называется блоком-35.
Различные G-факторы отвечают за узнавание различных групп промоторов и обеспечивают транскрипцию определенных генов (табл.1).
Таблица 4.1.
Последовательности, узнаваемые σ-факторами
|
σ-факторы E.coli |
Последовательность -35 |
Последовательность -10 |
|
σ70 |
TTGACA |
TATAAT |
|
σ32 |
TCTC-CCCTTGAA |
CCCCAT-TA |
|
σ54 |
(-24) CTGG-A |
(-12)TTGCA |
Инициация транскрипции включает образование двойного закрытого комплекса, затем формируется двойной открытый комплекс и после этого происходит синтез коротких олигорибо-нуклеотидов. После того, как синтезируется фрагмент РНК более 9 нуклеотидов, σ-фактор необратимо диссоциирует и транскрипция вступает в стадию элонгации. Элонгацию осуществляет кор-фермент - a2ßß’ (рис. 4.4).
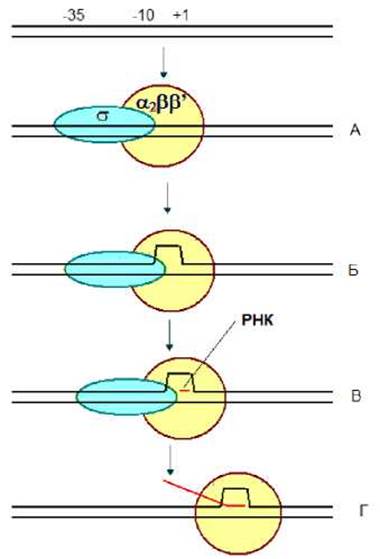
Рис. 4.4. Инициация транскрипции. А - закрытый двойной комплекс, Б - открытый двойной комплекс, В - открытый тройной комплекс, Г — элонгация транскрипции.
В процессе элонгации транскрипции образуется дуплекс РНК-ДНК, размер которого составляет около 12 пар нуклеотидов.
Кор-фермент способен синтезировать РНК со скоростью около 40 нуклеотидов в секунду.
При достижении терминатора кор-фермент завершает синтез РНК. У прокариот существует два типа терминаторов: р- зависимые и р-независимые. На р-зависимых терминаторах терминация осуществляется в присутствии белкового р-фактора. Такая терминация носит название р-зависимой терминации. Терминация транскрипции на независимых от р-фактора терминаторах называется р-независимой терминацией р-Независимая терминация обеспечивается образованием шпильки на РНК в процессе транскрипции и следующей за ней олигоуридиловой последовательностью. Шпилька приводит к паузе в транскрипции, а олигоуридил-олигоадениловый дуплекс, как наименее стабильный, диссоциирует во время паузы (рис. 4.5).
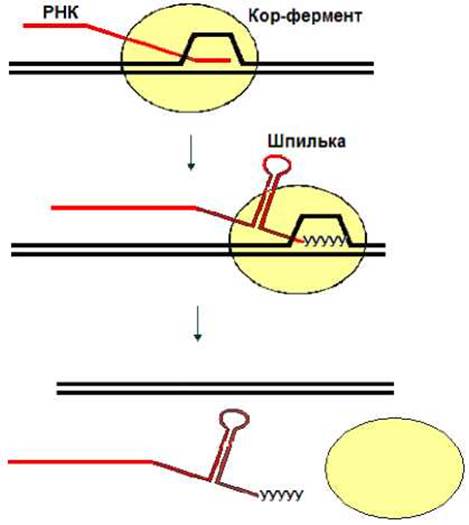
Рис. 4.5. р-Независимая терминация.
В случае р-зависимой терминации на синтезируемой РНК находится участок, с которым взаимодействует р-фактор (рис. 4.6). р-Фактор является НТФазой и способен использовать энергию гидролиза НТФ для движения по молекуле РНК от места посадки, расположенного в области 5’-конца РНК, в сторону ее 3’-конца. Как только р-фактор «догонит» работающую РНК-полимеразу, происходит терминация транскрипции (рис. 4.6).
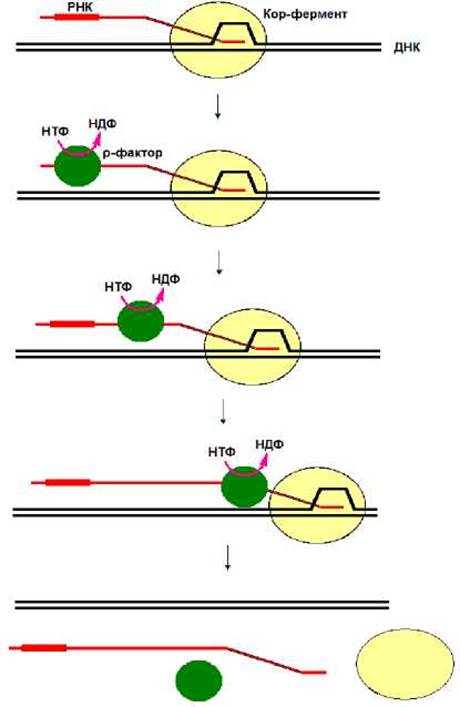
Рис. 4.6. р-Зависимая терминация.