Основы молекулярной биологии - В.И. Резяпкин 2009
Молекулярная организация генов
Ген - это элементарная единица наследственной информации, занимающая определенное место в геноме и ответственная за выполнение определенных функций в организме.
Гены представляют собой фрагменты НК, кодирующие полипептидные цепи или РНК, например рРНК, тРНК, мяРНК и др. Белки могут состоять из одной или нескольких полипептидных цепей, которые могут быть идентичными или отличающимися друг от друга. Если белок состоит из нескольких различных полипептидных цепей, то каждой цепи соответствует свой определенный ген (рис. 7.1).
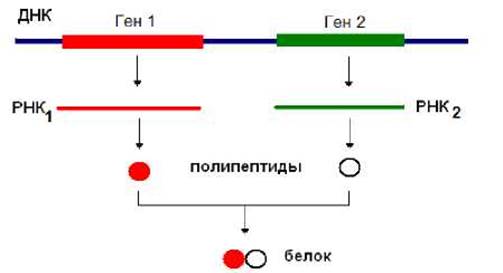
Рис. 7.1. Полипептидные цепи олигомерных белков могут быть закодированы в различных генах
В митохондриях есть белки, часть полипептидных цепей которых закодированы в ядерном геноме, другая часть в митохондриальном. Очевидно, что синтез полипептидных цепей, закодированных в ядерном геноме протекает в цитоплазме, других - в митохондриях. Полипептидные цепи, синтезированные в цитоплазме, транспортируются в митохондрии. Внутри этих органелл собирается нативный олигомерный белок (рис. 7.2).
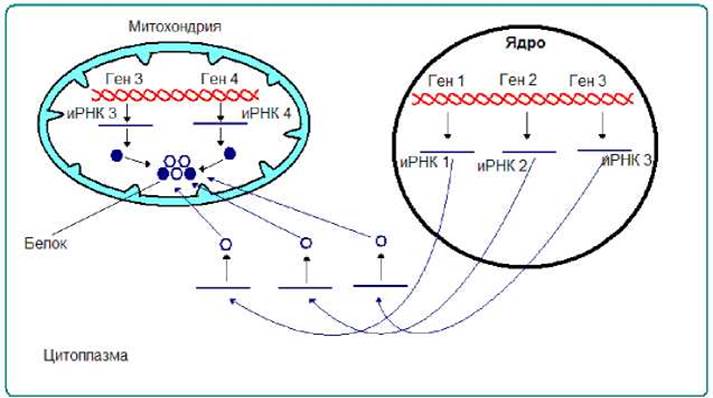
Рис. 7.2. Митохондриальные белки могут иметь смешанное происхождение
Гены могут быть уникальными - представленными в геноме одной копией и повторяющимися - представленными несколькими или множеством копий. Так гены гистонов присутствуют у некоторых видов эукариот в виде 1000 копий, гены рРНК также являются многократно повторяющимися. Более того, число генов рРНК в яйцеклетках амфибий может увеличиться в 3 раза за счет амплификации. Тем не менее, большинство генов эукариот в расчете на гаплоидный геном представлено одной копией или не большим числом копий. Прокариотические гены, как правило, присутствуют в геноме в виде одной копии.
Гены представленные большим числом копий могут быть разбросаны по геному или расположены рядом, образуя кластеры генов. В виде кластеров организованы гены гистонов, рРНК, глобинов - белков образующих гемоглобин - и др. Повторяющиеся гены расположенные последовательно друг за другом называются тандемными генами. Тандемный кластер генов образован повторяющимися единицами, которые в свою очередь состоят (рис. 7.3) из единицы транскрипции (транскрибируемый участок ДНК) и нетранскрибируемого спейсера (нетранскрибируемый участок ДНК).
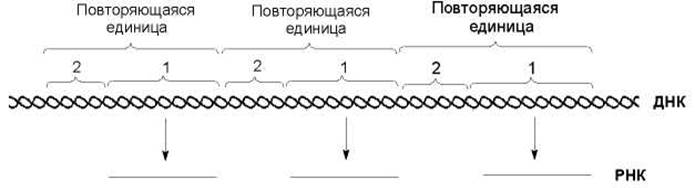
Рис. 7.3. Тандемный кластер генов образован повторяющими единицами, состоящими из единицы транскрипции (1) и нетранскрибируемого спейсера (2)
Как уже, отмечалось выше, гены гистонов имеют кластерную организацию. Различают пять основных гистонов Н1, Н2А, Н2В, Н3 и Н4. Эти белки являются родственными белками и выполняют общую функцию, связанную с поддержанием определенной структуры хроматина. Все пять генов гистонов входят в состав повторяющейся единицы, в результате последовательного расположения которых друг за другом формируется кластер. На рисунке 7.4 представлена повторяющаяся единица кластера гистоновых генов Drosophila melanogaster. У других видов эукариот повторяющаяся единица такого кластера организована иначе. В некоторых случаях гены гистонов не имеют кластерную организацию.
![]()
Рис. 7.4. Повторяющаяся единица кластера гистоновых генов Drosophila melanogaster. Стрелкой указано направление транскрипции генов
Гены располагаются на обеих цепях ДНК (рис. 7.5). Между ними обычно находятся некодирующие последовательности.
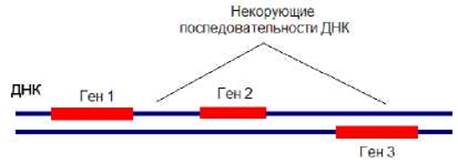
Рис. 7.5. Гены располагаются на обеих цепях ДНК
В то же время некоторые гены могут перекрываться, т.е. иметь общую последовательность ДНК. В одних случаях конец одного гена является началом другого (рис. 7.6А), в других - ген находится внутри другого гена (рис. 7.6В). Гены, расположенные на комплементарных цепях, также могут иметь общие участки (рис. 7.7). Перекрывание генов обнаружено, в частности, у вирусов. Данная особенность организации генов позволяет сэкономить ДНК, что особенно важно у вирусов, поскольку размеры их НК ограничены объемом вирусной частицы. Известны случаи, когда у вирусов на одном участке ДНК перекрывается три гена. В некоторых митохондриальных генах последний нуклеотид одного гена является первым нуклеотидом другого гена. Гены могут перекрываться со сдвигом или без сдвига рамки считывания. Если гены перекрываются без сдвига рамки считывания, то закодированные в них полипептиды будут иметь в области перекрывания идентичные аминокислотные последовательности. Если же гены перекрываются со сдвигом рамки считывания, то закодированные в них полипептиды не будут иметь идентичных аминокислотных последовательностей.
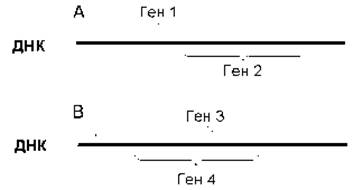
Рис. 7.6. Перекрывающиеся гены
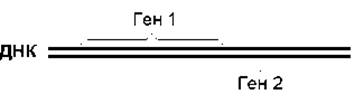
Рис. 7.7. Гены, расположенные на комплементарных цепях ДНК, могут иметь общие участки
Все гены можно разделить на две группы:
a) конститутивные гены, или «гены домашнего хозяйства». Эти гены постоянно экспрессируются: они функционируют на всех стадиях онтогенеза и во всех тканях. К ним относятся гены тРНК, рРНК, ДНК-полимеразы, РНК-полимеразы, рибосомальные белки, гистоны, гены, контролирующие постоянно протекающие обменные процессы;
b) индуцибельные гены или «гены роскоши», Эти гены могут включаться и выключаться. К индуцибельным генам относятся гены, контролирующие ход онтогенеза и гены, определяющие структуру и функции компонентов клетки и целостного организма. Включение индуцибельных генов называется индукцией, выключение - репрессией.
Гены могут подвергаться мутациям - изменениям последовательности нуклеотидов в цепи ДНК. Мутации могут возникать в результате замены, вставки или делеции (удаления) нуклеотидов. Мутации могут приводить к изменению аминокислотной последовательности закодированного в гене полипептида, а, следовательно, и к изменению биологических характеристик белка. Однако, благодаря эффекту вырожденности генетического кода не все изменения последовательности нуклеотидов приводят к изменению первичной структуры белка и, следовательно, к изменению его биологических функций.
Результатом мутации возможно перерождение генов в псевдогены. Псевдогены - это аналоги генов, неэкспрессирующиеся с образованием функционально активного продукта. Причинами превращения генов в псевдогены могут являться:
a) нарушение транскрипции;
b) нарушение созревания РНК;
c) нарушение трансляции;
d) другие причины.
Гены, как прокариот, так и эукариот состоят из регуляторной и транскрибируемой областей. Регуляторная область гена обеспечивает инициацию транскрипции транскрибируемой области гена. Последняя же несет информацию о полипептидной цепи или об определенных типах РНК (тРНК, рРНК и др.). Остановка транскрипции осуществляется в дистальной области гена - терминаторе.
У прокариот гены, кодирующие белки одного и того же метаболитического пути, могут быть сцеплеными и считываться с одного промотора в виде единой молекулы РНК, в результате трансляции которой образуется несколько полипептидов (рис.7.8).
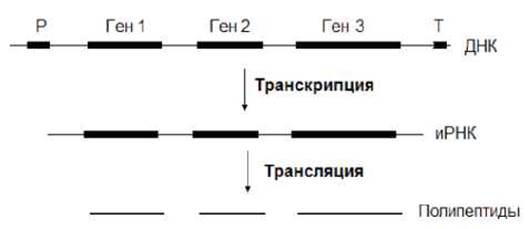
Рис. 7.8. У прокариот гены могут быть считаны с одного промотора в виде единой молекулы РНК, в результате трансляции которой образуется несколько полипептидов. Р - промотор, Т - терминатор.
Группа генов, имеющая общий промотор и терминатор, носит название оперон. На рисунке 7.9 представлена схема организации прокариотического оперона, кодирующего полипептиды. Регуляторная область такого оперона состоит из промотора - участка, с которым взаимодействует РНК-полимераза, и других нуклеотидных последовательностей, с которыми взаимодействуют регуляторные белки, обеспечивающие усиление или ослабление транскрипции. Транскрибируемая область состоит из кодирующих полипептидные цепи последовательностей и расположенными на их флангах нетранслируемых полинуклеотидных последовательностей. Оперонная организация генов обеспечивает согласованный синтез белков, участвующих в выполнении общей биологической функции.
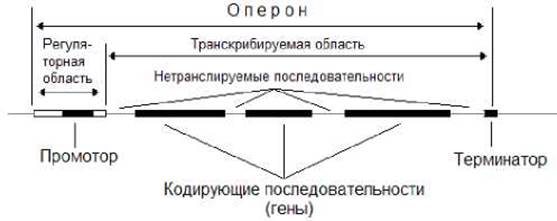
Рис. 7.9. Организация оперона
Гены, кодирующие белки, у прокариот могут быть также одиночными генами, состоящими из регуляторной и транскрибируемой областей (рис.7.10).
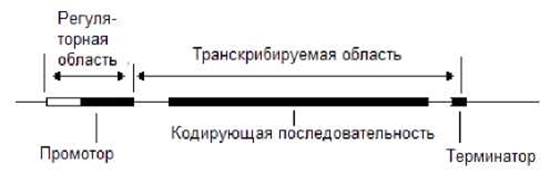
Рис. 7.10. Одиночный прокариотический ген
Гены рРНК прокариот имеют оперонную структуру. Интересно, что в состав таких оперонов входят гены тРНК (рис.7.11 А). Гены тРНК прокариот могут быть также представлены одиночными генами или объединены в составе оперона (рис.7.11 В).
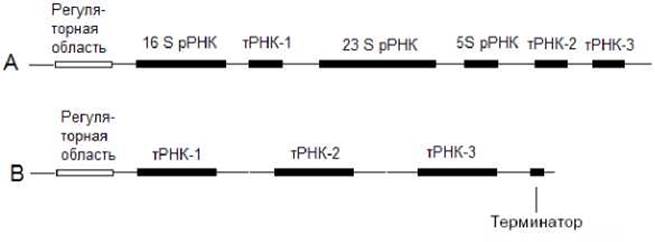
Рис. 7.11. Оперонная организация генов прокариот. А - оперон, содержащий гены рРНК и тРНК; В - оперон, содержащий гены тРНК
У эукариот гены устроены сложнее. Все гены эукариот можно разделить на три группы. Первая группа генов транскрибируется РНК-полимеразой I. К этой группе генов относятся гены 18S, 28S, 5,8S рРНК. Вторая группа генов транскрибируется РНК-полимеразой II. К этой группе генов относятся гены, кодирующие полипептиды и некоторые мяРНК. Третья группа генов транскрибируется РНК-полимеразой III. К этой группе генов относятся гены, кодирующие тРНК, 5S рРНК и гены других мяРНК. Гены всех трех групп отличаются друг от друга организацией промоторов. Именно это обстоятельство обеспечивает их транскрипцию различными РНК-полимеразами. Структура промоторов, узнаваемых различными РНК-полимеразами, была рассмотрена ранее в разделе «Транскрипция». В этом же разделе была рассмотрена организация генов рРНК и их экспрессия.
Еще одной особенностью РНК-полимераз эукариот является то, что они не могут самостоятельно инициировать транскрипцию. Для инициации транскрипции им необходима помощь белковых факторов - факторов транскрипции
На скорость транскрипции эукариотических генов оказывают влияние разнообразные регуляторные белки. Белки- активаторы усиливают транскрипцию, белки-ингибиторы оказывают тормозящие действие на экспрессию генов.
Далее более подробно рассмотрим организацию генов, транскрибируемых РНК-полимеразой II. Регуляторная зона таких генов состоит из промотора, на котором образуется комплекс РНК-полимеразы и общих факторов транскрипции и из регуляторных последовательностей (энхансеров и сайленсеров), с которыми связываются регуляторные белки. Известно шесть общих факторов транскрипции: TFIIA, TFIIB, TFIID, TFIIE, TFIIF, TFIIH. Они необходимы для инициации транскрипции. Одна из субъединиц TFIIH обладает протеинкиназной активностью, она осуществляет фосфорилирование (присоединение остатка фосфорной кислоты) РНК-полимеразы II. Фосфорилирование фермента необходимо для инициации транскрипции. Кроме общих факторов транскрипции в регуляции активности РНК-полимеразы II принимают участие и другие факторы транскрипции. В состав транскрипционного комплекса входят также белки, помогающие РНК-полимеразе разрушать нуклеосомы.
Первичные транскрипты эукариот гораздо длиннее иРНК. Причина этого заключается в том, что транскрибируемая область гена эукариот состоят из экзонов и интронов. При транскрипции таких генов синтезируется предшественник, в котором экзоны разделены интронами. В результате сплайсинга происходит удаление интронов и сшивка экзонов. Механизм сплайсинга и его варианты рассмотрены в разделе «Процессинг РНК».
Не все гены эукариот имеют экзон-интронную организацию. При этом доля генов с интронами в геноме различных видов неодинакова. С усложнением организации живых существ доля генов, состоящих из экзонов и интронов, возрастает, так у дрожжей таких генов 5 %, у дрозофилы - 83 %, у млекопитающих - 94 %.
В свою очередь варьируют количество и размеры интронов и экзонов (таблица). Так размер экзонов в среднем составляет 100-600 п.н. Размер же интронов варьирует в более широких пределах от нескольких десятков до десятков тысяч п.н. При этом общая длина интронов часто превышает общую длину экзонов. Например, в гене овальбумина кур из 7000 п.н. на долю экзонов приходится 1872 п.н.
Таблица 7.1
Характеристика экзонов и интронов некоторых генов
|
Ген |
организм |
Длина экзонов, пн |
Интроны |
|
|
число |
суммарная длина, пн |
|||
|
Ген а-интерферона |
человек |
600 |
0 |
0 |
|
Ген аденозиндезаминазы |
человек |
1500 |
11 |
30000 |
|
Ген аполипротеина B |
человек |
14000 |
28 |
29000 |
|
Ген ß-глобина |
мышь |
432 |
2 |
762 |
|
Ген а-глобина |
мышь |
463 |
2 |
256 |
|
Ген фиброина шелка |
шелкопряд |
18000 |
1 |
970 |
|
Ген гипоксантин- фосфорибозилтрансферазы |
мышь |
1307 |
8 |
32000 |
|
Ген эритропоэтина |
человек |
582 |
4 |
1562 |
|
Ген фазеолина |
фасоль |
1263 |
5 |
515 |
В связи с тем, что интроны удаляются в процессе сплайсинга РНК, кажется бессмысленным их существование. В то же время следующие сведения опровергают эту мысль. Удаление интронов в некоторых генах приводят к гибели организма. И еще один очень интересный факт. В пределах интрона может находиться другой ген. Этот ген в свою очередь может содержать свой интрон.
У инфузорий происходит удаление участков, соответствующих интронам, на уровне ДНК. Это явление осуществляется при созревании ДНК микронуклеуса. Интересно, что в процессе удаления фрагментов ДНК может происходить перетасовка участков, соответствующих экзонам (рис. 7.12).
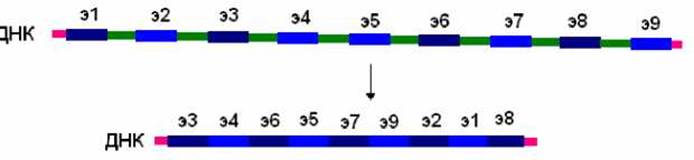
Рис. 7.12. У инфузорий происходит удаление участков, соответствующих интронам, на уровне ДНК. При этом может происходить перетасовка участков, соответствующих экзонам
Гены эукариот в отличие от прокариотических генов не организованы в опероны. В результате экспрессии эукариотических генов образуется моноцистронная иРНК, при трансляции которой образуется один полипептид.