Основы молекулярной биологии - В.И. Резяпкин 2009
Организация гемонов
Митохондриальный геном
Большинство митохондриальных геномов представляет собой двуцепочечные кольцевые суперспирализованные молекулы ДНК. У растений наряду с кольцевыми молекулами ДНК встречаются и линейные. У инфузорий, в митохондриях обнаружены только линейные молекулы ДНК.
Как правило, в клетке присутствуют ни одна митохондрия, при этом в ее составе содержится несколько ее геномов. Так, в клетках печени человека находится около 2 000 митохондрий, и в каждой из них содержится по 10 митохондриальных геномов, в клетках дрожжей S.cerevisiae содержится до 22 митохондрий, в которых по четыре генома, в фибробластах мыши - 500 митохондрий, по два генома в каждой.
В митохондриях многих организмов, за исключением высших животных, часть молекул ДНК находятся в виде олигомеров: линейных, кольцевых и цепных (катенаны), состоящих из продетых друг в друга, мономерных колец (рис.11.27).
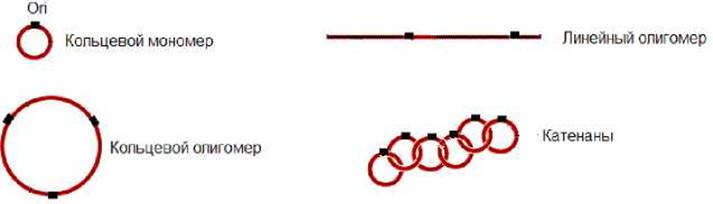
Рис. 11.27. В митохондриях ДНК могут находятся в виде мономеров и линейных, кольцевых и цепных (катенаны) олигомеров. Ori - точка начала репликации
Митохондриальный геном растений обычно состоит из нескольких кольцевых молекул ДНК разного размера. Одна из них большая и несколько молекул меньших размеров. Они могут между собой рекомбинировать. Так митохондриальный геном кукурузы может существовать в форме одной молекулы ДНК размером 570 000 п.н. или в виде нескольких молекул меньших размеров (рис. 11.28.). Существование митохондриального генома кукурузы в разных формах возможно благодаря процессам рекомбинации.
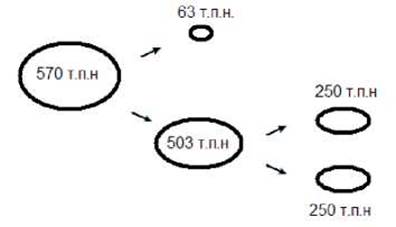
Рис. 11.28. Митохондриальный геном кукурузы может существовать в форме одной молекулы ДНК или в виде нескольких молекул меньших размеров
Размер митохондриального генома у разных организмов колеблется от нескольких тысяч до нескольких миллионов п. н. (таблица 11.6.). Размер минохондриальных геномов позвоночных обычно составляет от 16000 до 18000 п.н. Их ДНК компактна, большинство генов располагаются стык в стык; иногда они перекрываются на один нуклеотид, последний нуклеотид одного гена является первым нуклеотидом следующего. Наибольшую длину имеет митохондриальный геном растений. У некоторых видов он может достигать нескольких сотен тысяч и даже миллинов п.н. У растений, а также грибов и простейших митохондриальная ДНК в отличие от позвоночных содержит значительную долю, до 80 %, некодирующих последовательностей.
Таблица 11.6. Размер митохондриального генома
|
Организм |
Размер генома, п.н. |
|
Малярийный плазмодий |
∽6000 |
|
Простейшие |
22000 — 40000 |
|
Шпорцевая лягушка |
17533 |
|
Свинья |
16350 |
|
Человек |
16569 |
|
Дрожжи |
∽80000 |
|
Арабидопсис |
∽370000 |
|
Тыква |
∽850000 |
|
Дыня |
∽2400000 |
Состав и количество генов в митохондриальных геномах у разных видов значительно варьирует. У большинства высших животных геном митохондрий содержит 37 генов: 13 генов белков дыхательной цепи, 22 гена тРНК и два гена рРНК: 16S рРНК и 12S рРНК. На рисунке 11.29 представлен митохондриальный геном человека. У растений и простейших в митохондриальном геноме закодированы дополнительно и некоторые белки, входящие в состав рибосом. Наибольшее число генов, 97, обнаружено в митохондриальном геноме жгутикового простейшего Rectinomonas americana.
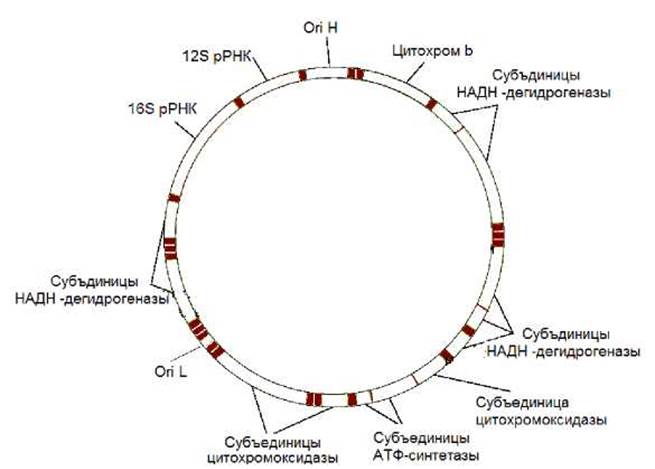
Рис. 11.29. Геном митохондрий человека. Его размер - 16569 п.н.. Содержит 13 генов, кодирующие полипептиды: 7 субъединиц НАДН-дегидрогеназного комплекса, 2 субъдиницы АТФ-синтетазы, 3 субъединицы цитохромоксидазы, 1 субъединицу убихинон-цитохром С редуктазы (цитохром b); 22 гена тРНК; 2 гена рРНК (16S РНК и 12S РНК). Ori H - точка начала репликации тяжелой цепи ДНК, Ori L - точка начала репликации легкой цепи ДНК.
Очевидно, что митохондриальный геном не обеспечивает синтез всех белков этих органелл, поскольку в организации и функционировании митохондрий принимают участие многие сотни белков. При этом большая их часть закодирована в ядерном геноме. Синтез таких белков происходит в цитоплазме. Затем они транспортируются в митохондрии, где и выполняют свои функции. К митохондриальным белкам ядерного происхождения относятся, многочисленные ферменты, переносчики электронов, транспортные белки, факторы, участвующие в транскрипции, трансляции и репликации митохондриальной ДНК и др. Интересно, что некоторые митохондриальные гены представлены копиями в ядерном геноме.
Одним из свойств кода является его универсальность, это означает, что у всех живых существ одни и те же триплеты кодируют одинаковые аминокислоты. При исследовании же генетического кода митохондрий оказалось, что он отличается от универсального. Более того, митохондриальные коды различных организмов не только отличаются от универсального, но и между собой. Наиболее изучен генетический код митохондрий позвоночных (рис. 11.30.). У них кодон АУА кодирует метионин вместо изолейцина в стандартном коде, кодоны АГА и АГГ, в стандартном коде кодирующие аргинин, являются стопкодонами, кодон УГА, в стандартном коде являющийся стопкодоном, кодирует триптофан. Таким образом, у животных в митохондриальном коде присутствуют коде 4 стоп-кодона и 60 смысловых. Интересно, что для прочтения всех 60 смысловых кодонов иРНК является достаточным 22 тРНК. В то время как для прочтения ядерного генетического кода необходимо как минимум 32 тРНК.
У иРНК митохондрий млекопитающих практически отсутствуют нетранслируемые 5- и 3’-концы. У них могут отсутствовать терминирующие кодоны. Такие иРНК оканчиваются на У или УА. При полиаденилировании их 3’-конца образуется терминирующий кодон УАА.
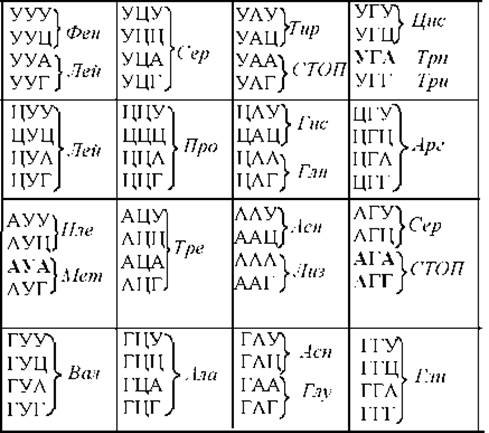
Рис. 11.30. Генетический код митохондрий позвоночных. Жирным выделены отличия от универсального ядерного кода.
Еще одной особенностью митохондриального генома многих организмов является редактирование РНК (см. раздел «Регуляция экспрессии генов»). Рассмотрим весьма любопытный пример редактирования митохондриальной иРНК. У Trypanosoma brucei редактируется иРНК одной из субъединиц цитохром-с-оксидазы посредством вставки четырех нуклеотидов У. Так образуется новая иРНК, служащая матрицей для синтеза новой субъединицы фермента, аминокислотная последовательность которой отличается от таковой, кодируемой нередактированной иРНК. Причиной синтеза субъединицы фермента с другой аминокислотной последовательностью является сдвиг рамки считывания, поскольку число вставленных нуклеотидов не кратно размеру триплета. Интересно, что новая субъединица образуется в митохондриях паразита, когда он попадает в организм холоднокровной мухи. Если же трипаносома живет в организме теплокровных млекопитающих, редактирования не происходит.
У большинства животных комплементарные цепи митохондриальной ДНК значительно отличаются по удельной плотности. В связи с этим одну из них обозначают как H-цепь (heavy - тяжелая), а другую - L-цепь (light - легкая). Каждая из цепей имеет свою точку начала репликации: репликация Н-цепи начинается в ori H, L-цепи - в ori L (рис. 11.31). Вначале происходит инициация репликация Н-цепи. Синтез же L-цепи начинается после того, как синтезируется значительная часть Н-цепи (у человека около 2/3 части всей длины тяжелой цепи). Таким образом, дочерние цепи митохондриальной ДНК синтезируются асинхронно.
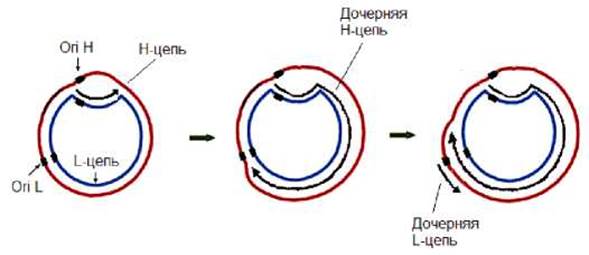
Рис. 11.31. Схема репликации митохондриальной ДНК млекопитающих
У млекопитающих транскрипция обеих цепей митохондрииальной ДНК начинается в районе ori H. В результате транскрипции образуются две длинные молекулы РНК, соответствующие по размеру Н- и L-цепям и комплементарны им. Кроме того, с Н-цепи считываются более короткие РНК, транскрипция которых останавливается на З'-конце гена 16S рРНК. (рис. 11.32). Коротких транскриптов образуется на порядок больше, чем длинных. В результате процессинга из коротких РНК образуются 12S рРНК и 16S рРНК, участвующие в формировании рибосом, а также две тРНК (тРНКфен и тРНКвал). Из длинных РНК вырезаются иРНК и остальные тРНК. На 3'-концах иРНК образуется поли-А-последовательность, как это характерно для эукариот. В то же время их 5'-концы не кэпируются. Поскольку гены млекопитающих не содержат интроны, их иРНК сплайсингу не подвергаются. Тем не менее, существуют митохондриальные геномы, содержащие гены с интронами. Например, в геноме митохондрий дрожжей присутствуют гены, имеющие интроэкзонную организацию. Митохондриальный геном дрожжей приблизительно в 5 раз больше такового человека и его размер составляет около 80 000 п.н. Однако, в нем содержится примерно столько же генов, как и в митохондриальной ДНК человека. Около 25 % от митохондриальной ДНК дрожжей приходится на АТ богатые участки, функции которых не совсем понятны.
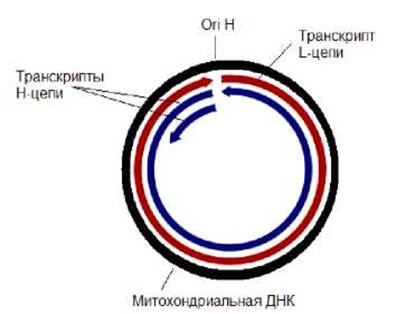
Рис. 11.32. Схема транскрипции митохондриальной ДНК млекопитающих Геном хлоропластов
Геном хлоропластов представляет собой кольцевую молекулу ДНК. Его размер находится в пределах от 130 до 200 тысяч п.н. В его составе представлено около 130 генов: по 2 гена рРНК (4,5S, 5S, 16S, 23S); около 30 генов тРНК; гены рибосомальных белков, РНК-полимеразы, около 40 генов белков тилакоидной мембраны и гены др. белков. Некоторые гены имеют интрон-экзонную организацию, другие являются непрерывными. Организация промоторов и терминаторов сходна с таковой прокариот. Интересно, что ДНК хлоропластов в отличие от ядерной ДНК может транскрибироваться РНК-полимеразой из Е. coli. Некоторые гены с близкими функциями (например, кодирующие белки рибосом) организованы сходным образом в геномах хлоропластов, Е. coli и цианобактерий
Не все белки хлоропластов закодированы в их геноме, большинство их имеет ядерное происхождение. Более того, полипептиды, кодируемые в хлоропластах, входят в состав ферментных комплексов, которые содержат субъединицы, кодируемых ядерным геномом. Не исключено, что в ходе эволюции эукариот значительная часть генов хлоропластов была перенесена в ядерный геном.
Аппарат белкового синтеза хлоропластов сходен с бактериальным, а не с эукариотическим. Их рибосомы имеют много общего с рибосомами Е. coli, сходны их полинуклеотидные последовательности. Рибосомы хлоропластов способны использовать тРНК прокариот при синтезе белка. Синтез белка в хлоропластах также как и у бактерий начинается с N- формилметионина, а не с метионина, как в цитоплазме клеток эукариот. иРНК хлоропластов эффективно транслируются белок-синтезирующим аппаратом Е. coli.
Существует гипотеза, что хлоропласты, как и митохондрии, произошли от прокариотических симбионтов, которые обитали в цитоплазме клеток эукариот. И этому мнению есть много подтверждений.