Общая микробиология - Шлегель Г. 1987
Основные механизмы обмена веществ и преобразования энергии
Важнейшие общие представления
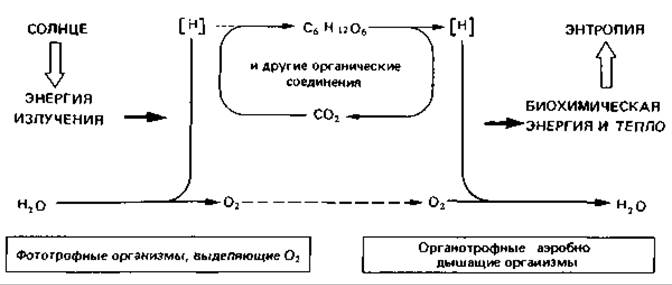
Рассматривая круговорот углерода, мы уже противопоставляли друг другу два процесса: фотосинтез, при котором фиксируется СO2 и выделяется О2, и минерализацию органических веществ, связанную с потреблением O2 и освобождением СO2. Процессы эти противоположны. С точки зрения массообмена важнее всего переход углерода из газообразного неорганического вещества в полутвердые и твердые органические соединения и обращение этого процесса. Если же рассматривать оба этих процесса с позиций энергообмена, т. е. преобразования энергии, то углерод имеет меньшее значение, чем водород. Еще Ю. Р. Майер (1848) сформулировал положение: «Растения поглощают одну силу - свет - и создают другую - химическое различие» («сила» здесь в смысле «энергия»), В процессе фотосинтеза лучистая энергия Солнца преобразуется в химическую энергию; вода при этом разлагается на кислород и водород, а последний в результате связывания с углеродом (из СО2) переводится в метастабильное состояние (см. схему). Большая часть такого стабилизированного водорода временно сохраняется в виде углеводов. Создаваемая растениями разность потенциалов между водородом и кислородом служит источником энергии для всех дышащих кислородом живых существ. В их организмах водород снова высвобождается из соединения с углеродом и соединяется с кислородом в «биохимической реакции гремучего газа» с выделением энергии. Это окисление водорода - многоступенчатый процесс, в ходе которого освобождающаяся энергия может отдельными порциями превращаться в биохимическую энергию. В глобальном масштабе система «фототрофные растения - органотрофные живые существа» включается в процесс преобразования энергии излучения в тепло и приводит к замедлению роста энтропии.
Метаболизм и метаболические пути. Как во время роста, так и в состоянии покоя вегетативные клетки нуждаются в постоянном притоке энергии. Живая клетка представляет собой высокоорганизованную материю. Энергия необходима не только для создания такой организации, но и для ее поддержания. Эту энергию организм получает в процессе обмена веществ, или метаболизма, т. е. путем регулируемых превращений, которым различные вещества подвергаются внутри клеток. Источниками энергии служат питательные вещества, поступающие из внешней среды. В клетках эти вещества претерпевают ряд изменений в результате последовательных ферментативных реакций, образующих этапы определенных метаболических путей. Такие пути выполняют две главные функции: они, во-первых, поставляют материалы-предшественники для построения клеточных компонентов и, во-вторых, обеспечивают энергию для клеточных синтезов и других процессов, требующих затраты энергии.
Превращения веществ в клетке (обмен веществ, или метаболизм), в результате которых из сравнительно простых предшественников, например глюкозы, жирных кислот с длинной цепью или ароматических соединений, образуется новое клеточное вещество, можно ради простоты подразделить на три основные группы. Сначала питательные вещества расщепляются на небольшие фрагменты (распад, или катаболизм), а затем в ходе реакций промежуточного обмена, или амфиболизма, они превращаются в ряд органических кислот и фосфорных эфиров. Эти два пути переходят незаметно один в другой. Многообразные низкомолекулярные соединения - это тот субстрат, из которого синтезируются основные строительные блоки клетки. «Строительными блоками» мы называем аминокислоты, пуриновые и пиримидиновые основания, фосфорилированные сахара, органические кислоты и другие метаболиты — конечные продукты цепей биосинтеза, иногда длинных. Из них строятся полимерные макромолекулы (нуклеиновые кислоты, белки, резервные вещества, компоненты клеточной стенки и т. п.), из которых состоит клетка. Эти два этапа биосинтеза клеточных веществ - синтез строительных блоков и синтез полимеров - составляют синтетическую ветвь метаболизма, или анаболизм (рис. 7.1).
Биохимическое единство. Принцип «биохимического единства» - одна из немногих догм, признаваемых в нашем веке. Согласно этому принципу, все живые существа, обитающие на Земле, в биохимическом отношении в основе своей сходны. Этот принцип выражается, например, в единообразии строительных блоков (в том числе в однотипности их оптического вращения), во всеобщей роли аденозинтрифосфата (АТР) как элементарного кванта биологической энергии, в универсальности генетического кода, а также в единстве путей превращения сахаров и природы дыхательной цепи. Почти идентичны у всех живых существ и главные метаболические пути. Существует лишь несколько групп бактерий, у которых основные схемы метаболизма так или иначе модифицированы - преобладают какие-то определенные пути, а другие укорочены или иным образом изменены. Все варианты обмена веществ микроорганизмов без труда можно свести к общей схеме. Метаболические пути складывались, очевидно, в ходе эволюции, и можно думать, что типичный для аэробных организмов биохимический аппарат возник сравнительно поздно, когда в воздухе появился кислород. В настоящее время трудно решить, что представляют собой укороченные метаболические пути - примитивные особенности или результат деградации.
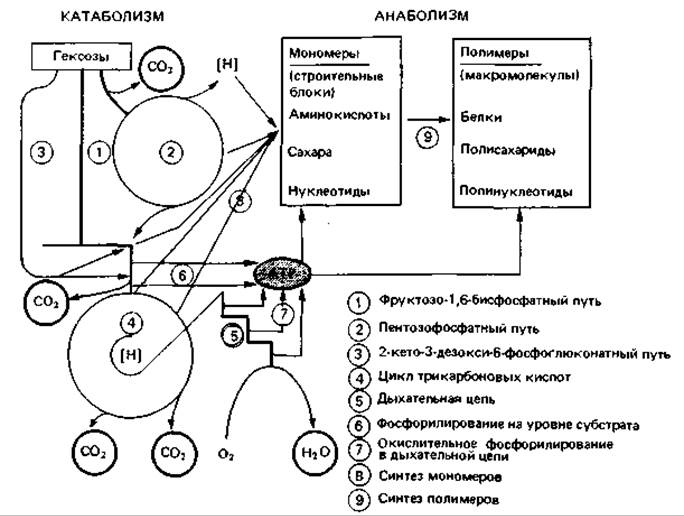
Рис. 7.1. Схема обмена веществ у аэробно дышащих клеток, расщепляющих гексозы.
Катаболизм углеводов. Как уже отмечалось при описании круговорота углерода, углеводы - главный продукт фотосинтеза у растений. В то же время для большинства микроорганизмов они служат основными питательными веществами. Поэтому в дальнейшем при рассмотрении питательных веществ - субстратов клеточного метаболизма - мы будем в первую очередь говорить о глюкозе. Что касается других природных веществ, используемых микроорганизмами в качестве субстратов, то о них речь пойдет позже в связи с другими явлениями (гл. 14). Макромолекулы, как правило, сначала расщепляются вне клетки выделяемыми ею ферментами (экзоферментами) на мономерные и димерные блоки и лишь в такой форме поглощаются клеткой.
Гексозы после ряда предварительных этапов расщепляются «пополам». Продукты расщепления превращаются в пировиноградную кислоту (пируват), которая занимает ключевое положение в промежуточном метаболизме, так как служит исходным соединением во многих процессах синтеза и распада. В результате декарбоксилирования пирувата образуются С2-соединения, которые связываются сначала с подходящей акцепторной молекулой (оксалоацетатом), а затем в цикле трикарбоновых кислот, называемом также циклом лимонной кислоты, постепенно окисляются до СО2. Оксалоацетат (щавелевоуксусная кислота) в этом циклическом процессе регенерируется. Атомы водорода (или восстановительные эквиваленты), отщепившиеся на разных этапах окисления органических веществ, поступают в ATP-регенерирующую систему дыхательной цепи (окислительное фосфорилирование). При каждом обороте цикла трикарбоновых кислот из одного Отсоединения (ацетил-кофермента А) образуются две молекулы СО2 и четыре раза по 2[Н]. Эти реакции выравнивают баланс цикла трикарбоновых кислот.
К промежуточным соединениям цикла трикарбоновых кислот относятся органические кислоты, поставляющие исходный материал для процессов биосинтеза (2-оксоглутарат, сукцинат, оксалоацетат). Таким образом, цикл трикарбоновых кислот не только участвует в конечном окислении питательных веществ, но служит также важным «распределителем», поставляющим исходные соединения для синтеза основных структурных единиц («строительных блоков») клетки. Если бы указанные кислоты постоянно выводились из цикла, то регенерации молекулы-акцептора не происходило бы и цикл был бы нарушен. Так называемые анаплеротические последовательности реакций обеспечивают поступление в цикл трикарбоновых кислот все новых количеств промежуточных соединений взамен израсходованных для биосинтеза. Эти анаплеротические последовательности имеют особо важное значение для тех организмов, которые растут за счет простых одно- или двухуглеродных соединений или других субстратов, разлагающихся на такие же простые соединения.
Роль ферментов. Химические превращения осуществляются в клетке с помощью ферментов. За каждое превращение одного метаболита в другой ответствен особый фермент. Ферменты представляют собой белки, обладающие каталитической функцией. Основные свойства ферментного белка заключаются в его способности распознавать определенные метаболиты, катализировать их превращения и обеспечивать регуляцию каталитической активности.
Катализируемая ферментом реакция начинается со связывания определенного метаболита (субстрата) с ферментным белком. Каждый фермент взаимодействует, как правило, лишь с одним метаболитом - своим субстратом - и катализирует его превращение в другой метаболит до установления равновесия. Таким образом, каждый фермент характеризуется определенной субстратной специфичностью (взаимодействует только с одним метаболитом и продуктом его превращения) и определенной специфичностью действия (катализирует лишь одно из многочисленных превращений, которым может подвергаться данный метаболит). Узнавание субстрата ферментом происходит в процессе связывания. Субстрат присоединяется в совершенно определенном участке молекулы ферментного белка - так называемом каталитическом центре. Стерические свойства субстрата и распределение зарядов в его молекуле служат теми признаками, по которым субстрат распознается ферментом. Субстрат и фермент подходят друг к другу, как ключ к замку.
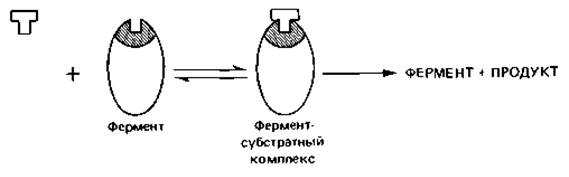
Ферментные белки действуют как биокатализаторы, снижая энергию активации. Химические превращения метаболита на ферменте протекают при обычной температуре. Ферменты, таким образом, обеспечивают протекание таких реакций, которые без них могли бы происходить только при высокой температуре или при других нефизиологических условиях, которые клетка не могла бы выдержать.
Скорость реакции, катализируемой ферментом, примерно на 10 порядков выше, чем скорость неферментативной реакции; увеличение скорости в 1010 раз сокращает полупериод какой-либо реакции с 300 лет до одной секунды.
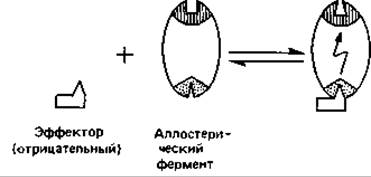
Очень важное свойство ферментов, окончательно установленное лишь сравнительно недавно, состоит в том, что их каталитическая активность подвержена регуляции. Эта регулируемость ферментной активности - одно из возможных объяснений гармоничного протекания всех метаболических процессов в клетке. По крайней мере некоторые ферменты (хотя бы по одному в каждом специфическом пути биосинтеза) подвергаются регуляторным воздействиям. Такие ферменты с помощью своего каталитического центра распознают субстрат, а с помощью другого центра - конечный продукт данной цепи реакций или иные низкомолекулярные вещества, определенным образом влияющие на их активность. У этих ферментов имеется второй связывающий участок - регуляторный центр. Связывание конечных продуктов или других метаболитов, называемых также эффекторами, влияет на каталитический центр, изменяя его активность. Конечные продукты действуют как отрицательные эффекторы. Положительные эффекторы повышают активность фермента. Таким образом, концентрации метаболитов, играющих роль эффекторов, определяют активность фермента, а тем самым и скорость соответствующих превращений. Эффекторы по своей структуре не имеют ничего общего с субстратами ферментов. Они стерически отличны от субстратов. Поэтому говорят об аллостерических эффекторах, а центры, ответственные за регуляцию, называют аллостерическими центрами ферментов.
Коферменты и простетические группы. В связывании и последующем переносе отдельных фрагментов субстрата, например водорода, метальных групп, аминогрупп и т. п., наряду с ферментными белками участвуют низкомолекулярные соединения - так называемые коферменты и простетические группы (табл. 7.1, рис. 7.2). Коферменты и простетические группы более или менее прочно связаны с ферментами. Вещества, которые на ферментном белке присоединяют к себе фрагмент субстрата, а затем отделяются от него, чтобы передать этот фрагмент на другом ферментном белке второму соединению, называют коферментами (более точным было бы название косубстраты или переносчики). Низкомолекулярные соединения, прочно связанные с белковой частью фермента и не отделяющиеся от нее во время присоединения и переноса фрагментов субстрата, называются простетическими группами этих ферментов.
Таблица 7.1. Коферменты и простетические группы, функционирующие как переносчики водорода, различных групп или электронов, и отношение их к витаминам
|
Кофермент или простетическая группа1 |
Функция (что подвергается переносу) |
Витамин |
|
|
NAD (Р) |
(221) |
Водород, е- |
Никотиновая кислота |
|
FMN |
(237) |
Водород, е- |
Рибофлавин |
|
FAD |
(219) |
Водород, е- |
Рибофлавин |
|
Убихинон |
(237) |
Водород, е- |
|
|
Цитохромы |
(237) |
е- |
Производные гема |
|
Биотин |
Карбоксильные группы |
Биотин |
|
|
Пиридоксальфосфат |
(432) |
Аминогруппы |
Пиридоксин |
|
Тетрагидрофолиевая кислота |
(301) |
Формильные группы |
Фолиевая кислота, 4-аминобензойная кислота |
|
Кофермент А |
(219) |
Ацильные группы |
Пантотеновая кислота |
|
Липоевая кислота |
(219) |
Ацильные группы и водород |
Липоевая кислота |
|
Тиаминпирофосфат |
(219) |
Альдегидные группы |
Тиамин |
|
Кофермент В12 |
Карбоксильные группы (перемещение внутри молекулы); метильные группы |
Кобаламин |
|
1 Цифры в скобках указывают, на какой странице приводится химическая формула.
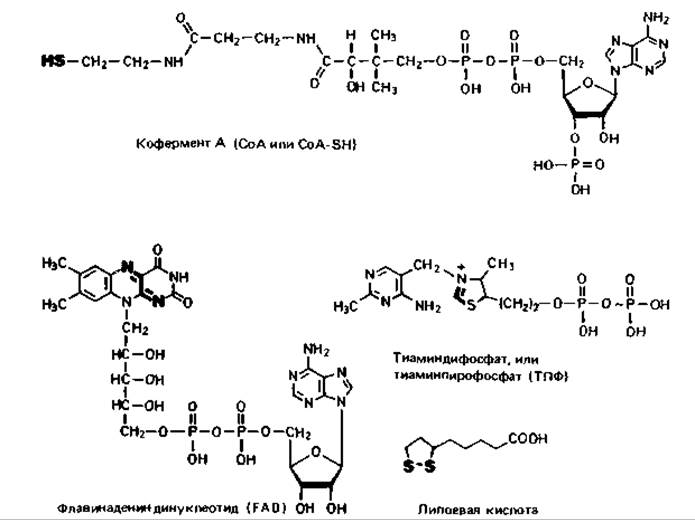
Рис. 7.2. Структурные формулы некоторых коферментов и простетических групп. Активные группы выделены жирным шрифтом.
Коферменты имеют особое значение, так как многие организмы не способны их синтезировать и должны получать с пищей в виде витаминов. Многим молочнокислым бактериям, почвенным и водным бактериям, а также другим одноклеточным организмам для роста необходимы те или иные из витаминов, приведенных в табл. 6.2, или их предшественники, которые добавляют в питательную среду.
Преобразование энергии. Кратко обрисованные выше пути метаболизма (превращения глюкозы, цикл трикарбоновых кислот, дыхательная цепь) приводят к окислению сахара до СО2 и воды. При этом освобождается столько же энергии, сколько при сжигании сахара; но благодаря тому, что окисление глюкозы разбито на ряд отдельных ферментативных реакций, теоретически полностью обратимых, выделяющаяся при окислении энергия может переводиться в биохимически доступную форму без значительного повышения температуры.
Во многих реакциях освобождаются лишь незначительные количества энергии. Такие минимальные выходы энергии полезны для клетки лишь в тех случаях, когда равновесие реакции сдвинуто в сторону образования продуктов. При некоторых превращениях, связанных с выделением большого количества свободной энергии (- ∆G = 40-60 кДж/моль, или 10-15 ккал/моль), эта энергия запасается в процессе фосфорилирования на уровне субстрата в форме АТР и может затем использоваться в реакциях, требующих затраты энергии. В такой регенерации АТР участвуют как субстраты (промежуточные соединения), так и ферменты. Однако большая часть энергии, выделяющейся в результате окисления питательных веществ, переводится в доступную для клетки форму (в высокоэнергетические связи АТР) в процессе окислительного фосфорилирования в электрон-транспортной (дыхательной) цепи.
Метаболиты. Уже при поверхностном знакомстве с веществами, участвующими в клеточном метаболизме, легко заметить, что многие из них находятся в фосфорилированном состоянии, т.е. в форме эфиров фосфорной кислоты. Нефосфорилированные промежуточные продукты содержат карбоксильные группы или ионизующиеся основные группы. Создается впечатление, что ферменты могут воздействовать только на те метаболиты, в которых имеется ионизированная, т.е. заряженная, группа. Незаряженные молекулы или группы всегда связаны с коферментами или простетическими группами ферментов; некоторые образуют шиффовы основания с диаминокислотой лизином, содержащейся в активном центре ферментного белка. Неионизированными бывают лишь соединения, стоящие в начале и в конце тех или иных метаболических путей; таковы многие субстраты и некоторые выделяемые клеткой продукты обмена (глюкоза, фруктоза, этанол, ацетон, 2-пропанол, бутанол, глицерол и т.д.). Остается пока открытым вопрос о том, связано ли наличие ионизированных промежуточных соединений с функциями ферментов или с особой способностью клетки удерживать такие метаболиты.
Дегидрирование и пиридиннуклеотиды. Окисление органических соединений происходит путем переноса электронов от донора к акцептору. При биологическом окислении субстрата чаще всего происходит одновременный перенос двух электронов; при этом от субстрата отщепляются также два протона (Н +). Такое окисление субстрата, формально происходящее с отщеплением двух атомов водорода, называется дегидрированием. Нередко термины донор водорода и донор электронов употребляются как синонимы; равнозначны и такие термины, как акцептор водорода и акцептор электронов; окисление и дегидрирование; восстановление и гидрирование.
Ферментные белки, отщепляющие от субстратов атомы водорода, называются дегидрогеназами, причем в названиях некоторых из них содержится указание на донор водорода (лактатдегидрогеназа, малатдегидрогеназа и т.д.). Многие дегидрогеназы переносят водород на один из двух коферментов - никотинамидадениндинуклеотид (NAD) или никотинамидадениндинуклеотидфосфат (NADP).
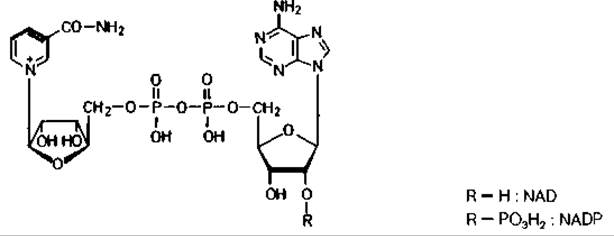
Группой, определяющей функцию обоих коферментов, является амид никотиновой кислоты. Один водород переносится с субстрата вместе с парой электронов (в виде гидрид-иона) на пиридиновое кольцо, второй переходит в раствор. Этот перенос стереоспецифичен: одни ферменты (алкогольдегидрогеназа, лактатдегидрогеназа) переносят водород на одну сторону пиридинового кольца, а другие (глицеральдегидфосфат-дегидрогеназа) - на другую сторону. Для краткости такое обратимое дегидрирование изображают следующим образом:
![]()
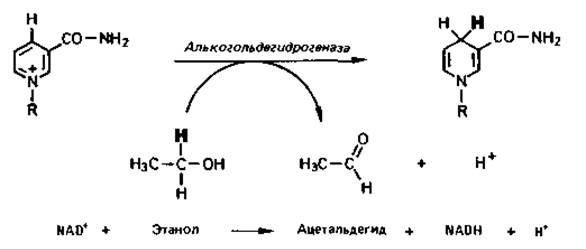
У восстановленных форм обоих коферментов в отличие от окисленных форм максимум поглощения лежит при 340 нм. Поэтому восстановление и окисление коферментов можно проследить по изменению поглощения в соответствующей области спектра. На этом основаны многие оптические методы определения активности ферментов.
Оба кофермента свободно диссоциируют, т. е. отделяются от одного белка-дегидрогеназы и переносят водород после связывания с другой дегидрогеназой на другой акцептор. Поэтому их называют также переносчиками водорода. NADH2 переносит водород в основном на предшественники конечных продуктов брожения или же передает его в дыхательную цепь, в то время как NADPH2 участвует главным образом в восстановительных этапах процессов биосинтеза.
ATP и другие высокоэнергетические соединения. Процессы, требующие затраты энергии, могут осуществляться в клетке благодаря аденозинтрифосфату (АТР). В форме АТР энергия, полученная в результате фотосинтеза, дыхания или брожения, становится доступной для клетки и может быть ею использована. АТР - универсальный переносчик химической энергии между реакциями, доставляющими энергию, и реакциями, потребляющими ее (это вещество называют «энергетической валютой» клетки). АТР служит непосредственным источником энергии для таких разнообразных процессов, как синтез структурных компонентов макромолекул, механическое движение и осмо регуляция. Пирофосфатные связи между фосфатными группами «богаты энергией». Это означает, что у них высокий потенциал переноса групп. Для образования этих связей требуется больше энергии, чем для образования обычной эфирной связи; соответственно и при их разрыве тоже выделяется много энергии (при гидролизе изменение свободной энергии AG'0% — 30 кДж) или же она запасается в продуктах реакции (табл. 7.2).
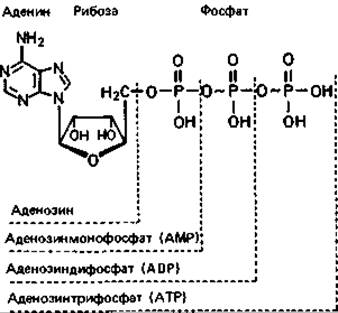
Таблица 7.2. «Богатые» и «бедные» энергией соединения, имеющие биохимическое значение. Указана свободная энергия — ∆G'0гидролиза при pH 7,0 в стандартных условиях
|
Субстрат |
-∆G'0 |
|
|
кДж |
ккал |
|
|
Ацетилфосфат |
44,0 |
10,5 |
|
Ацетоацетил-СоА |
44,0 |
10,5 |
|
Ацил-АМР |
55,7 |
13,3 |
|
Креатинфосфат |
37,7 |
9,0 |
|
Фосфоенолпируват |
54,4 |
13,0 |
|
Простые фосфорные эфиры |
12,6 |
3,0 |
|
Гликозиды |
12,6 |
3,0 |
|
Сахароза |
27,6 |
6,6 |
|
UDP-глюкоза |
31,8 |
7,6 |
|
Альдозо-1-фосфат |
20,9 |
5,0 |
|
АТР (→ ADP + Pi) |
31,0 |
7,4 |
|
АТР (→ АМР + РРi) |
31,8 |
7,6 |
АТР как кофермент для активации метаболитов. Многие промежуточные соединения нуждаются в активации путем переноса групп. При этом возможны три способа расщепления АТР:
1. Сахара активируются путем превращения в соответствующие фосфорилированные производные:
Глюкоза + АТР → Глюкозофосфат + ADP
2. Рибозо-5-фосфат активируется в результате переноса дифосфатного остатка (пирофосфата):
Рибозо-5-фосфат + АТР → Фосфорибозилдифосфат + АМР
3. Некоторые из неорганических кислот, все аминокислоты и неорганический сульфат активируются путем присоединения АМР-группы с освобождением пирофосфата:
Жирная кислота + АТР → Ацил-АМР + Пирофосфат
Аминокислота + АТР → Аминоацил-МАР + Пирофосфат
Сульфат + АТР → Сульфат-АМР + Пирофосфат
Доведение реакции до конца (т.е. завершение такого превращения) обеспечивается тем, что образующийся пирофосфат расщепляется пирофосфатазой и тем самым выводится из сферы реакции. Таким образом, активация этих метаболитов обходится клетке в две высокоэнергетические связи.
Приведенных сведений, вероятно, достаточно для того, чтобы ясно показать универсальное значение АТР, а также подвести читателя к пониманию различных типов метаболизма у бактерий. Знакомясь с этими типами, следует иметь в виду, что клетка, используя в данных условиях доступные ей питательные вещества, всегда стремится получить возможно большее количество АТР.
Синтез (регенерация) АТР осуществляется в основном с помощью трех процессов: фотосинтетического фосфорилирования (разд. 12.2), окислительного фосфорилирования (фосфорилирование в дыхательной цепи, разд. 7.4) и фосфорилирования на уровне субстрата (разд. 7.2.1). Два первых процесса сходны между собой в том, что АТР образуется в них при участии ATP-синтазы. Субстратное фосфорилирование может происходить при различных реакциях промежуточного метаболизма. В обмене углеводов важнейшие реакции, приводящие к регенерации АТР, катализируются фосфоглицераткиназой, пируваткиназой и ацетаткиназой. Бактерии и дрожжи, сбраживающие сахара, располагают лишь тем АТР, который образуется с помощью этих ферментов. Во всех таких процессах фосфорилирования (за редкими исключениями) акцептором фосфата служит аденозиндифосфат (ADP). Аденозинмонофосфат (АМР), прежде чем он будет фосфорилирован в АТР, должен превратиться в ADP в реакции, катализируемой аденилаткиназой (АМР + ATP → 2ADP).
Ниже будут кратко описаны четыре важнейших этапа катаболизма глюкозы-начальное расщепление ее до С3-соединений, цикл трикарбоновых кислот, окисление пирувата и дыхательная цепь, а также функции этих процессов. При этом мы остановимся на некоторых подробностях, важных для понимания ряда специализированных метаболических процессов. Для более глубокого ознакомления с физиологией обмена веществ и биохимией микроорганизмов необходимо обратиться к учебникам по биохимии.