Общая микробиология - Шлегель Г. 1987
Типы брожения
Cпиртовое брожение, вызываемое дрожжами и бактериями
Этиловый спирт (этанол) - один из широко распространенных продуктов сбраживания сахаров микроорганизмами. Даже растения и многие грибы в анаэробных условиях накапливают этанол. Главные продуценты этанола - дрожжи, особенно штаммы Saccharomyces cerevisiae. Дрожжи, как и большинство других грибов, осуществляют аэробное дыхание, но без доступа воздуха они сбраживают углеводы до этанола и СО2. У ряда анаэробных и факультативно-анаэробных бактерий этиловый спирт тоже является главным или побочным продуктом сбраживания гексоз или пентоз.
8.1.1 Образование этанола дрожжами
В изучении описанных выше путей обмена важную роль сыграли работы, проведенные на дрожжах. Еще Гей-Люссак (в 1815 г.) дал уравнение, описывающее превращение глюкозы в этанол в той форме, как это принято и теперь:
![]()
Нормальное сбраживание глюкозы дрожжами. Сбраживание глюкозы до этанола и СO2 дрожжами (Saccharomyces cerevisiae) осуществляется по фруктозобисфосфатному пути. Превращение пирувата в этанол происходит в два этапа. Сначала пируват декарбоксилируется пируватдекарбоксилазой (а) при участии тиаминпирофосфата до ацетальдегида, а затем ацетальдегид восстанавливается алкогольдегидрогеназой (б) в этанол при участии NADH2:
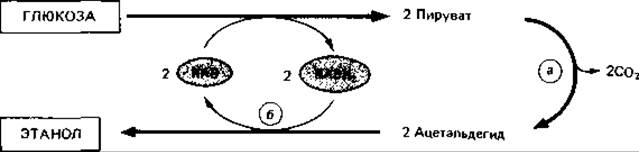
Переносится при этом водород, образующийся при дегидрировании триозофосфата; окислительно-восстановительный баланс, таким образом, сохраняется.
История исследования дрожжевого брожения. Нет смысла останавливаться здесь на курьезно звучащих в наше время спорах о природе брожения - о том, является ли образование спирта из сахара результатом «химического контактного действия» или же результатом деятельности живых существ. Л. Пастер окончательно разрешил вопрос о причинах брожения и установил, что дрожжи в аэробных условиях образуют из определенного количества сахара примерно в 20 раз больше клеточного вещества, чем в анаэробных. Он открыл также, что кислород подавляет брожение. Этот эффект, получивший название эффекта Пастера, приобрел с тех пор известность как один из классических примеров регуляции обмена веществ.
В 1896-1897 годах Бухнер и Хан обнаружили, что если к соку, полученному при растирании прессованных пивных дрожжей с кварцевым песком и кизельгуром, добавить сахар, то начинается брожение с образованием пены. Так впервые сложный биохимический процесс удалось осуществить вне клетки. В 1906 году Гарден и Ионг, тоже работавшие с дрожжевым соком, установили, что для сбраживания глюкозы нужен неорганический фосфат, который включается в фруктозо-1,6-бисфосфат. Сбраживание глюкозы дрожжевым соком происходит согласно уравнению Гардена-Ионга:
![]()
Формы брожения, открытые Нейбергом. Открытия Карла Нейберга и разработанные им оригинальные методы имеют не только историческое значение. Этот исследователь показал, что дрожжи способны сбраживать помимо глюкозы также и пируват. В качестве промежуточного продукта при сбраживании пирувата образуется ацетальдегид; это можно доказать, связывая альдегид бисульфитом (который для дрожжей практически нетоксичен). Если к дрожжам, сбраживающим глюкозу, прибавить бисульфит, будет происходить следующая реакция:
![]()
При этом появится новый продукт брожения - глицерол - и одновременно снизится выход этанола и СО2.
Брожение в присутствии бисульфита стали использовать в промышленности при производстве глицерола. Эта технология основана на том, что ацетальдегид связывается и поэтому не может служить акцептором водорода. Вместо ацетальдегида роль такого акцептора принимает на себя дигидроксиацетонфосфат; он восстанавливается до глицерол-3-фосфата и дефосфорилируется с образованием глицерола. Брожение можно представить так:
![]()
Это модифицированное дрожжевое брожение известно как вторая форма брожения по Нейбергу. Принцип перехватывания одного из метаболитов - метод ловушки - стал впоследствии общим биохимическим методом.
При добавлении к бродящему раствору NaHCO3 или Na2HPО4 тоже образуется глицерол, так как ацетальдегид превращается в результате реакции дисмутации в этанол и уксусную кислоту и поэтому не может быть использован в качестве акцептора водорода. Это третья форма брожения по Нейбергу:
![]()
Нормальное дрожжевое брожение Нейберг назвал первой формой брожения. Он считал, что образующийся при небиологическом разложении фруктозы 2-оксопропаноль (метилглиоксаль, СН3-СО-СНО) также является промежуточным продуктом сбраживания глюкозы.
Отношение дрожжей к кислороду. Сбраживание дрожжами глюкозы-анаэробный процесс, хотя дрожжи - аэробные организмы. В анаэробных условиях брожение идет очень интенсивно, но роста дрожжей почти не происходит. При аэрации брожение ослабеваем уступая место дыханию. У некоторых дрожжей можно почти полностью подавить брожение усиленной аэрацией (эффект Пастера). Пастер открыл этот эффект более ста лет тому назад, исследуя процессы брожения при изготовлении вина. Это явление свойственно не только дрожжам, но и всем другим факультативно-анаэробным клеткам, включая клетки тканей высших животных.
Баланс превращений глюкозы (на примере типичного опыта с дрожжами) представлен в табл. 8.1. Аэрация уменьшает потребление глюкозы, а также образование этанола и СО2, но делает возможным рост дрожжей. С энергетической точки зрения эти явления понятны; они указывают на существование у дрожжей чрезвычайно полезного регуляторного механизма: в анаэробных условиях образуются только 2 моля АТР на один моль использованной глюкозы, а при дыхании - 38 молей АТР. Таким образом, клетка, регулируя превращения субстрата, может получать максимум энергии как в тех, так и в других условиях.
Таблица 8.1. Баланс превращений глюкозы при анаэробной и аэробной инкубации дрожжей (при 25°С) с 2,4-динитрофенолом (ДНФ) и без него. Числа указывают количество глюкозы, потребленной, расщепленной или ассимилированной в единицу времени. (Lynen F., Netter К. J., Schuegraf А. und G. Stix. «Phosphatkreislauf und Pasteur-Effekt. 8. Colloquium d. Ges f. Physiol. Chemie, Mosbach, Springer-Verl., Berlin, 1958)
|
Анаэробные условия |
Аэробные условия |
Аэробные условия + 0,4 мМ ДНФ |
|
|
Потребление |
- 3,085 |
- 1,950 |
- 3,020 |
|
Расщепление |
- 1,945 |
- 0,699 |
- 1,943 |
|
Брожение |
- 1,945 |
- 0,343 |
- 1,387 |
|
Дыхание |
— |
- 0,356 |
- 0,556 |
|
Ассимиляция |
- 1,145 |
- 1,251 |
- 1,077 |
В третьем столбце табл. 8.1 приводятся результаты опыта, в котором дрожжи инкубировали в аэробных условиях в присутствии 0,4 мМ 2,4-динитрофенола (ДНФ). ДНФ разобщает фосфорилирование и окисление в дыхательной цепи: он нарушает сопряжение между переносом электронов и фосфорилированием, после чего дыхание протекает уже без контроля со стороны фосфорилирования. Добавление ДНФ практически выключает фосфорилирование в дыхательной цепи, и водород, отщепляющийся в цикле трикарбоновых кислот, уже не может использоваться для энергетических целей. Доступным для использования оказывается только высокоэнергетический фосфат, образующийся при расщеплении сукцинил-СоА. Результаты опыта показывают, что в присутствии ДНФ потребление глюкозы возрастает до величины, наблюдаемой в анаэробных условиях, что связано с его разобщающим действием.
В реакции Пастера участвует, видимо, несколько регуляторных механизмов, действующих в одном направлении. Эффект одного из них проявляется на уровне процессов фосфорилирования. В основе его лежит конкуренция за аденозиндифосфат (ADP) и неорганический фосфат (Рі). Для дегидрирования при расщеплении глицеральдегидфосфата необходимы ортофосфат и ADP:
![]()
Таким образом, расщепление субстрата (глюкозы) по фруктозобисфосфатному пути зависит от наличия ADP и неорганического фосфата. В отсутствие ADP и фосфата дегидрирование глицеральдегидфосфата оказывается невозможным. Однако в аэробных условиях с этой реакцией конкурирует за ADP и фосфат процесс фосфорилирования в дыхательной цепи, который тоже приводит к образованию АТР. Вполне вероятно, что расщепление глюкозы, а вместе с тем и образование этанола тормозятся в результате снижения внутриклеточной концентрации ADP и фосфата. Если же под действием ДНФ процессы окисления в дыхательной цепи и фосфорилирование разобщаются, то ADP и фосфат снова могут использоваться для дегидрирования глицеральдегидфосфата, и аэробное потребление глюкозы повышается до уровня, соответствующего анаэробным условиям (табл. 8.1).
За реакцию Пастера ответствен еще один регуляторный механизм, а именно аллостерический эффект торможения фермента фосфофруктокиназы аденозинтрифосфатом. Это торможение будет рассмотрено в разделе 16.2.2.
Объяснение процесса, описанного Гарденом и Ионгом. Отжатый дрожжевой сок разлагает глюкозу в соответствии с уравнением Гардена - Ионга. Накопление фруктозо-1,6-бисфосфата объясняется в этом случае тем, что АТР в нарушенной ферментной системе (в отличие от живой клетки) не может использоваться для энергетических целей и остается в избытке. Дрожжевой сок не содержит фосфатаз, поэтому ADP должен непрерывно регенерироваться за счет фосфорилирования лишней глюкозы или фруктозо-6-фосфата.
Применение дрожжей. Человек использует дрожжи в различных целях. Уже в древние времена из множества видов, рас и штаммов были отобраны наиболее подходящие для того или другого процесса. Для производства пива применяются слабо дышащие, преимущественно бродящие формы, относящиеся к так называемым «низовым дрожжам». Для получения спирта и вин и в хлебопечении используются главным образом «верховые дрожжи».
Пекарские дрожжи (Saccharomyces cerevisiae) должны поднимать тесто в результате образования СО2, т. е. осуществлять интенсивное брожение. Их выращивают в чанах при сильной аэрации. В качестве побочного продукта при этом всегда образуется этанол. Варьируя степень аэрации и режим добавления сахара, можно изменять относительный выход дрожжей и спирта. При проточном способе непрерывно прибавляют сахар, но так медленно, чтобы он ограничивал рост дрожжей. Это избавляет от образования продуктов брожения - весь сахар используется для роста. Источником азота служит аммоний. Необходимые ростовые вещества дрожжи получают из добавляемого пшеничного затора.
Пивные дрожжи по большей части относятся к низовым расам (мюнхенское, пильзенское пиво), реже к верховым (светлое пиво, эль, портер). В Центральной Европе пиво варят в основном из ячменя, выбирая для этой цели ячмень с наименьшим содержанием белка и высоким содержанием крахмала. Поскольку дрожжи не образуют амилаз и могут сбраживать только сахара, но не крахмал, необходимо сначала обеспечить осахаривание крахмала. Для этого используют специфическую амилазу, образующуюся при прорастании зерен ячменя. Зернам дают набухнуть и прорасти; образовавшийся зеленый солод осторожно сушат при определенной температуре, при которой прерывается только процесс прорастания, а ферменты сохраняются. Просушенный солод размалывают и погружают в чаны с водой. При умеренной температуре крахмал через некоторое время осахаривается, превращаясь в мальтозу. Полученное сусло освобождается от дробины, к нему добавляют хмель, варят, охлаждают и подвергают брожению в бродильных чанах, прибавляя предварительно выращенные дрожжи.
Для получения спирта (этанола) путем брожения используют отходы производства сахара (мелассу) или картофель. Большие количества дешевого спирта получают также из гидролизатов древесины лиственных пород или из сульфитных щелоков -отходов бумажных фабрик. В гидролизатах древесины сбраживаются до этанола только гексозы. Остающиеся после этого пентозы используют для выращивания кормовых дрожжей (Endomyces lactis и видов Torula), для которых они служат источником углерода; эти дрожжи, богатые белками, добавляют в корм скоту.
Многие немецкие вина получают в результате спонтанного брожения виноградного сока, вызываемого дрожжами рода Kloeckera. Для подавления неконтролируемого брожения, вызываемого дикими дрожжами, неоднократно вносят чистые культуры дрожжей или смеси Kloeckera и Saccharomyces, которые подбирают в специальных винодельческих районах. Еще большее значение для букета вина имеет, однако, сорт винограда (Рислинг, Сильванер, Мюллер-Тургау, Шейребе, Кернер и др.), а также климатические и почвенные факторы, влияющие на рост виноградной лозы.
Во всех жидкостях, полученных путем дрожжевого брожения, содержатся сивушные масла: пропанол, 2-бутанол, 2-метилпропанол, амиловый (пентанол) и изоамиловый (триметилбутанол) спирты. Они представляют собой продукты нормального бродильного метаболизма дрожжей и обнаруживаются не только при их росте в сложных питательных растворах, содержащих аминокислоты. Основными компонентами сивушного масла являются побочные продукты обмена изолейцина, лейцина и валина.
8.1.2 Образование этанола бактериями
Известный для дрожжей путь образования этанола (фруктозобисфосфатный путь, пируватдекарбоксилазная реакция) из всех исследованных бактерий обнаружен только у Sarcina ventriculi. Из пульке - бродящего сока агавы (Agave mexicana) в Мексике была выделена палочковидная подвижная бактерия с полярными жгутиками, образующая этанол. Эта бактерия Zymomonas mobilis разлагает глюкозу по 2-кето-3-дезокси-6-фосфоглюконатному пути и расщепляет пируват с помощью пируватдекарбоксилазы на ацетальдегид и двуокись углерода. Ацетальдегид восстанавливается до этанола. Единственными продуктами этого брожения являются этанол, СО2 и небольшие количества молочной кислоты. Интересно, что в водке, приготовленной из сока агавы, обнаруживаются углеродные атомы 2, 3, 5 и 6 глюкозы, тогда как в спирте, образуемом дрожжами, содержатся углеродные атомы 1, 2, 5 и 6 (рис. 8.1).
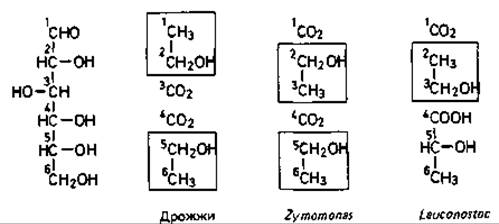
Рис. 8.1. Происхождение углеродных атомов этанола из углеродного скелета глюкозы при сбраживании глюкозы.
При брожениях, вызываемых некоторыми видами Enterobacteriaceae и клостридиями, этанол является побочным продуктом. Предшественник этанола - ацетальдегид - образуется в этом случае не прямо из пирувата (под действием пируватдекарбоксилазы), а путем восстановления ацетил-СоА.
Совершенно иным путем образуют этиловый спирт гетероферментативные молочнокислые бактерии (например, Leuconostoc mesenteroides). Глюкрза разлагается ими по пентозофосфатному пути до пентозофосфата. На ксилулозо-5-фосфат воздействует фосфокетолаза:
![]()
Образовавшийся ацетилфосфат восстанавливается ацетальдегид-дегидрогеназой и алкогольдегидрогеназой в этанол. Другой продукт расщепления глюкозы - глицеральдегид-3-фосфат - восстанавливается до лактата (через пируват).