Общая микробиология - Шлегель Г. 1987
Перенос электронов в анаэробных условиях
Образование сероводорода при восстановлении сульфата
Физиологическую группу бактерий, восстанавливающих сульфат (их называют также десульфатирующими, сульфатредуцирующими или сульфидобразующими), отличает способность к переносу водорода с субстрата на сульфат как конечный акцептор электронов и, таким образом, к восстановлению сульфата до сульфида. В этом процессе происходит перенос электронов, и в нем участвует цитохром с. Энергия запасается благодаря фосфорилированию в электрон-транспортной цепи в анаэробных условиях:
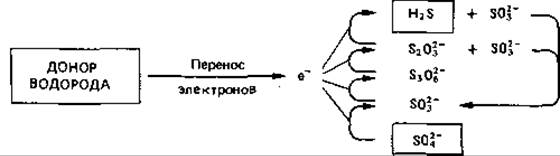
Поскольку такое восстановление сульфата обладает формальным сходством с дыханием, при котором акцептором водорода служит кислород, принято говорить о сульфатном дыхании, или о диссимиляционной сулъфатредукции. Главным продуктом такого процесса является сероводород:
![]()
Большая часть сероводорода, образующегося в природе, возникает благодаря этой реакции. Сульфатредуцирующие бактерии являются, в отличие от нитратредуцирующих, облигатными анаэробами, т. е. нуждаются в строго анаэробных условиях.
Систематика. Сульфатредуцирующие бактерии - это физиологическая группа, для которой характерна способность к образованию сероводорода из сульфата (табл. 9.1). Донорами водорода служат простые низкомолекулярные соединения, образующиеся при анаэробном разложении биомассы, главным образом целлюлозы: лактат, ацетат, пропионат, бутират, формиат, этанол, высшие жирные кислоты и молекулярный водород. По степени усвоения органических кислот различают две группы сульфатредуцирующих бактерий :
1. Бактерии, относящиеся к первой группе, окисляют донор водорода не полностью и выделяют уксусную кислоту. Таковы виды спорообразующего рода Desulfotomaculwn (D. nigrificans, D. orientis и D. ruminis) и неспорообразующего рода Desulfovibrio (D. vulgaris, D. desulfuricans, D. gigas, D. thermophilus и другие).
2. Вторая группа включает роды и виды, часть которых может расти, используя спирты, ацетат, высшие жирные кислоты или бензоат, а другие способны даже к хемоавтотрофному росту в присутствии водорода и формиата. К этой группе относятся спорообразователи (Desulfotomaculwn acetoxidans), а также неспорообразующие палочки (Desulfobacter), кокки (Desulfococcus), сарцины (Desulfosarcina), нитевидные формы, передвйгающиеся путем скольжения (Desulfonema), и некоторые другие бактерии.
Таблица 9.1. Важнейшие и наиболее распространенные сульфатредуцирующие бактерии
|
Рост на средах различного состава |
||||||||
|
Вид |
Форма клеток |
Жгутикование |
Цитохром (характерный) |
Десульфовиридин |
с пируватом (без сульфата) |
с малатом (и сульфатом) |
с формиатом (и сульфатом) |
Термо- фильность |
|
Desulfovibrio desulfuricans |
Вибрионы |
Полярное, монотрихальное |
с3 |
+ |
+ |
+ |
- |
- |
|
с3 |
+ |
- |
- |
- |
- |
|||
|
D. vulgaris D. gigas |
Спириллы |
Полярное, политрихальное |
с3 |
+ |
- |
- |
- |
|
|
Desulfotomaculum nigrificans |
Палочки |
Перитрихальное |
b |
- |
+ |
+ |
||
|
Dm. orientis |
Изогнутые палочки |
» |
b |
- |
- |
- |
- |
|
|
Dm. ruminis |
Палочки |
» |
b |
+ |
+ |
- |
||
|
Dm. acetoxidans |
» |
Полярное |
b |
- |
- |
- |
- |
- |
Восстановление сульфата. Почти все бактерии, грибы и зеленые растения способны использовать в качестве источника серы сульфат. Они получают сульфид, необходимый для синтеза серусодержащих аминокислот, путем «ассимиляционной сульфатредукции». Первая реакция на этом пути является общей как для диссимиляционного, так и для ассимиляционного восстановления сульфата. Далее при диссимиляционной сульфатредукции происходит прямое восстановление активированного сульфата, а при ассимиляционной следует еще одна реакция активации. Восстановление сульфата в клетке начинается с его активации, на которую непосредственно затрачивается энергия АТР (рис. 9.3); с помощью АТР-сульфурилазы (сульфатаденилтрансферазы) дифосфатный остаток АТР обменивается на сульфат:
![]()
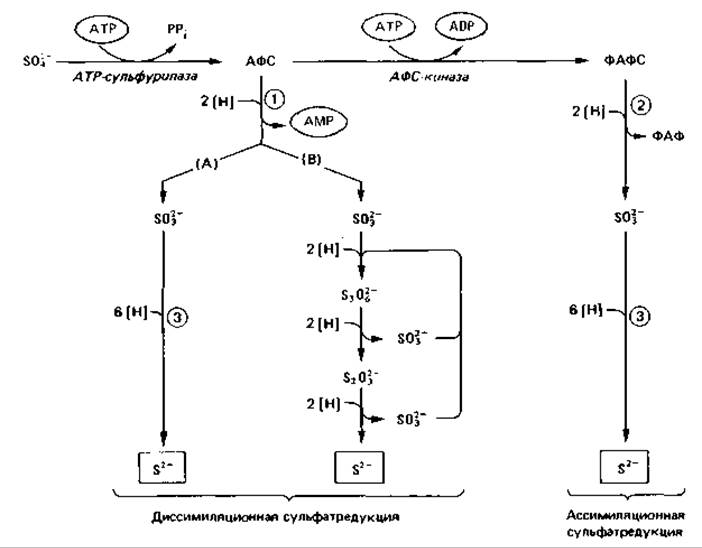
Рис. 9.3. Схема диссимиляциоиного восстановления сульфата («сульфатного дыхания») и ассимиляционного восстановления сульфата. АФС - аденозин-5'-фосфосульфат; ФАФС - фосфоаденозин-5'-фосфосульфат; ФАФ - фосфоаденозин-5'-фосфат. Ферменты, участвующие в реакциях (цифры в кружках): 1 - АФС-редуктаза; 2 - ФАФС-редуктаза; 3 - сульфитредуктаза (бисульфитредуктаза).
Дифосфат (пирофосфат) расщепляется пирофосфатазой. Продуктом активации является аденозин-5-фосфосульфат (АФС). Последующие реакции могут быть различными. На пути ассимиляционного восстановления сульфата АФС с помощью АФС-киназы и АТР фосфорилируется у ряда организмов с образованием фосфоаденозинфосфосульфата (ФАФС); лишь этот вдвойне активированный сульфат восстанавливается сначала до сульфита, а затем до сульфида. При диссимиляционной сульфатредукции АФС с помощью АФС-редуктазы восстанавливается до сульфита, что сопровождается образованием АМР.
Восстановление сульфита до сульфида происходит, видимо, у разных бактерий по-разному. С помощью сульфитредуктазы сульфит прямо восстанавливается до сульфида (на что затрачивается 6 электронов) без образования промежуточных продуктов. В такого рода восстановлении - как и в ассимиляционной сульфитредукции - участвуют, по-видимому, железопорфириновые соединения (десульфовиридин, десульфорубидин). Второй механизм состоит в последовательном трехступенчатом восстановлении сульфита с образованием промежуточных продуктов, таких как тритионат и тиосульфат (рис. 9.3). Предполагается, что электроны для восстановления сульфита доставляются цитохромами (у одних бактерий-цитохромом b, у других - цитохромом с).
Фосфорилирование, сопряженное с переносом электронов. Предположение о такого рода фосфорилировании у сульфатредуцирующих бактерий основано на данных о наличии цитохромов и железосерных белков в плазматических мембранах, а также о высоком выходе энергии. Цитохром с3 обладает, по сравнению с другими цитохромами, весьма низким окислительно-восстановительным потенциалом (Е'0 = - 205 мВ) и находится на внешней поверхности мембраны или в периплазматическом пространстве.
У хорошо изученных видов сульфатредуцирующих бактерий была обнаружена конститутивная гидрогеназа (Н2 : цитохром-с3-оксидоредуктаза), с помощью которой Н2 может как поглощаться и активироваться, так и выделяться в окружающую среду. Некоторые сульфатредуцирующие бактерии растут в присутствии Н2 и сульфата как единственных источников энергии. Способность к восстановлению сульфата с помощью Н2 и к образованию больших количеств сероводорода, не связанному с заметным ростом, вероятно, характерна для большинства сульфатредуцирующих бактерий.
Перенос электронов с Н2 как донора, сопровождающийся восстановлением 1 моля сульфата до 1 моля сульфида, вероятно, сопряжено с регенерацией 3 молей АТР, из которых, однако, 2 моля расходуются на активацию сульфата.
Окисление органических субстратов. Классические сульфатредуцирующие бактерии, которые были известны до 1975 года (например, Desulfovibrio vulgaris), окисляют органические субстраты не до Н2O и СO2, а до уксусной кислоты. Эти бактерии не имеют полного цикла трикарбоновых кислот. Однако недавно было выделено несколько видов, способных окислять ацетат, высшие жирные кислоты и бензоат.
Ассимиляция органических субстратов. Энергия, получаемая сульфатредуцирующими бактериями в результате окислительного фосфорилирования, делает возможной ассимиляцию органических веществ (органических кислот, аминокислот и т. п.). Некоторые штаммы способны синтезировать клеточные компоненты из ацетата и СO2, если донором водорода служит Н2. Организмы, ассимилирующие органические вещества в процессе окисления неорганического донора электронов, можно называть хемолитогетеротрофами. Фиксация СO2 в цикле Кальвина у данной группы микроорганизмов не обнаружена.
Брожение без сульфата. Некоторые сульфатредуцирующие бактерии обладают способностью расщеплять лактат или пируват в отсутствие
сульфата. Вместо окисления пирувата
![]()
они осуществляют брожение с выделением Н2:
![]()
Таким образом, сульфатредуцирующие бактерии можно отнести к микроорганизмам, осуществляющим брожение.
Получение накопительных культур и выделение. Для выделения сульфатредуцирующих бактерий необходимо использовать питательную среду, содержащую подходящий донор водорода, пригодный для ассимиляции углеродный субстрат, минеральные вещества и сульфат; необходимо поддерживать анаэробные условия и достаточно низкий окислительно-восстановительный потенциал (E'0= — 200 мВ) (рис. 9.4).
Распространение и роль сульфатредуцирующих бактерий в природе. Сульфатредуцирующие бактерии встречаются главным образом в сероводородном иле, где органические вещества подвергаются анаэробному разложению. Эти бактерии, по-видимому, специально приспособлены к использованию продуктов неполного разложения углеводов - таких веществ, как жирные кислоты, гидроксикислоты, спирты и водород. Основную массу образующегося в природе сероводорода следует считать конечным продуктом сульфатного дыхания. Загрязненные воды содержат от 104 до 106 сульфатредуцирующих бактерий на 1 мл, а сероводородный ил - до 107.
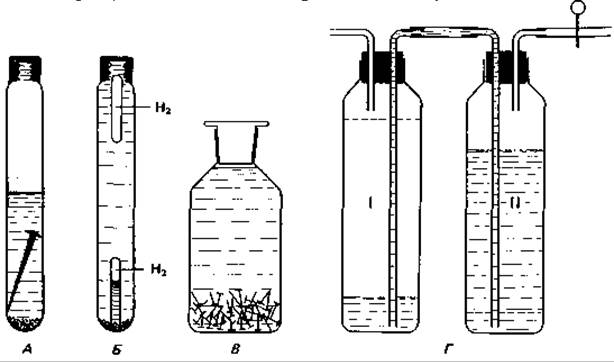
Рис. 9.4. Накопительные культуры сульфатредуцирующих бактерий. Рост после инокуляции среды сероводородным илом. А. Среда содержит лактат и сульфат; железный гвоздь обеспечивает достаточно низкий окислительно-восстановительный потенциал (в результате катодной поляризации). Б. Доказательство использования молекулярного водорода сульфатредуцируюхцими бактериями: пробирка Дёрхема, заполненная Н2, перед инкубацией плавает, а после инкубации засеянной среды оказывается на дне. В. Рост бактерий в закупоренной бутыли в присутствии малых количеств органического вещества за счет восстановления сульфата и анаэробной коррозии железа. Г. Накопление сульфатредуцирующих бактерий в двойном сосуде Зёнгена. Сосуд II заполняют средой, содержащей лактат и сульфат; затем впускают в него Н2, и часть жидкости переходит в сосуд I (новые уровни показаны пунктирными линиями); в течение двухдневной [Инкубации при 30°С значительная часть Н2 потребляется.
Большинство разрабатываемых месторождений серы (например, в Техасе, Луизиане и Мексике) имеет невулканическое происхождение; это отложения биогенной серы прошлых геологических эпох. Путем восстановления сульфатов морской воды за счет органических отходов (сточных вод) с помощью сульфатредуцирующих бактерий можно получать сероводород, а следовательно, и серу.
Большое экономическое значение имеет косвенный результат жизнедеятельности сульфатредуцирующих бактерий (например, Desulfovibrio) - анаэробная коррозия железа. Во влажной среде ионизация железа может происходить и в анаэробных условиях:
(1) Окисление железа:
![]()
Обычно образующаяся при этом пленка из молекулярного водорода предохраняет железо от дальнейшего разрушения. Однако в присутствии сульфатредуцирующих бактерий и при наличии в среде сульфатов происходит катодная деполяризация, и тогда железо окисляется даже в отсутствие кислорода;
(2) Восстановление сульфата:
![]()
(3) Образование сульфида железа (выпадающего в осадок):
![]()
Суммарная реакция (1-3):
![]()
Обусловленное такой коррозией повреждение железных труб весьма убыточно.
Способность сульфатредуцирующих бактерий использовать органические кислоты, спирты и даже молекулярный водород, образующийся при поляризации железа, в качестве доноров водорода лежит в основе способа получения накопительных культур этих бактерий (рис. 9.4).
Сульфатредуцирующие бактерии считают ответственными за высокое содержание сероводорода в глубинных слоях Черного моря (глубже 200 м). В Венеции гондолы красят обычно в черный цвет; по-видимому, это защитная мера, связанная с тем, что красители, содержащие тяжелые металлы, изменяют под влиянием H2S свой первоначальный цвет.
В образовании сероводорода в рубце жвачных животных участвует Desulfotomaculum ruminis (разд. 14.1).