Общая микробиология - Шлегель Г. 1987
Фототрофные бактерии и фотосинтез
Пурпурные и зеленые бактерии
Способность использовать свет как источник энергии, необходимой для роста, присуща двум группам бактерий, принципиально отличающимся друг от друга.
Пурпурные и зеленые бактерии, объединяемые в порядок Rhodospirillales1, можно рассматривать как реликтовые организмы, дошедшие до нас из времен начальной эволюции фотосинтеза. Они не в состоянии использовать в качестве донора водорода воду (как это делают зеленые растения); им требуются доноры с более высокой степенью восстановления (H2S, H2 или органические вещества). Поэтому фотосинтез у этих бактерий протекает без выделения O2. В таких случаях говорят об аноксигенном фотосинтезе. Бактерии этой группы - типичные водные организмы, распространенные как в пресной, так и в морской воде. Их красная, оранжевая или зеленая окраска обусловлена присутствием бактериохлорофиллов и каротиноидов.
Цианобактерии используют в качестве донора водорода воду и выделяют на свету кислород. Таким образом, они осуществляют оксигенный фотосинтез. Пигментная система этих бактерий включает хлорофилл а, каротиноиды и фикобилины. Поскольку процесс фотосинтеза у цианобактерий принципиально не отличается от фотосинтеза зеленых растений, эту группу бактерий до недавнего времени рассматривали совместно с фотосинтезирующими эукариотами и называли сине-зелеными водорослями. Однако по строению своих клеток это типичные прокариоты. Цианобактерии уже были подробно описаны в разделе 3.21 и здесь рассматриваться не будут2.
1 В настоящее время зеленые бактерии относят к особому порядку Chlorobiales, а вместе с пурпурными бактериями их объединяют в класс Anoxyphotobacteria. - Прим. ред.
2 Кроме цианобактерий к оксигенному фотосинтезу способны прокариотические организмы, составляющие порядок Prochlorales. Вместе с цианобактериями их относят к классу Oxyphotobacteria. - Прим. ред.
Фототрофные бактерии, осуществляющие аноксигенный фотосинтез, делятся на две большие группы: пурпурные бактерии (Rhodospirillales) и зеленые бактерии (Chlorobiales). Представители этих двух порядков значительно различаются по своим цитологическим и физиологическим признакам, а также по характерным для них пигментам (табл. 12.1; рис. 12.1, 12.6 и 12.10).
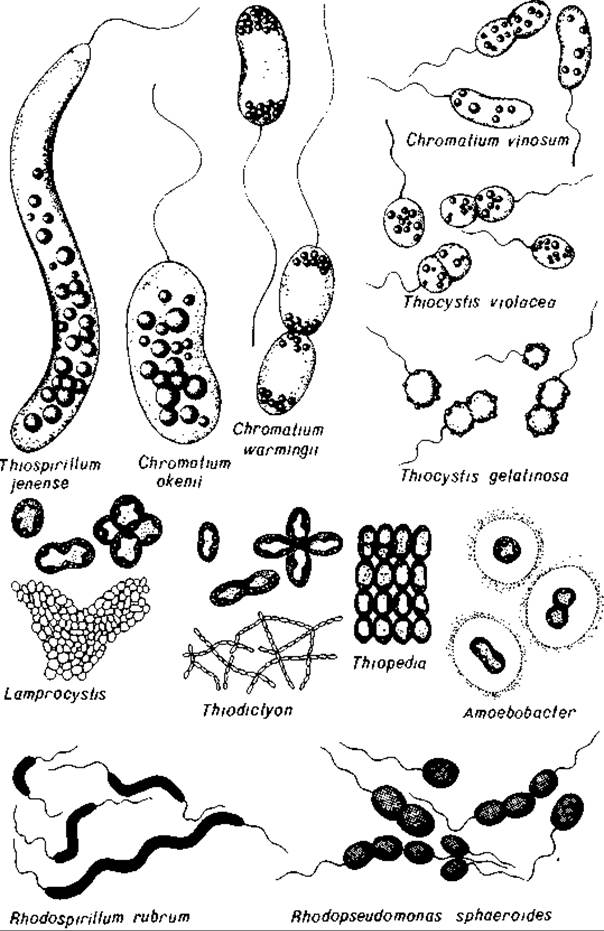
Рис. 12.1. Некоторые представители серных пурпурных бактерий (Chromatiaceae) и несерных пурпурных бактерий (Rhodospirillaceae).
Пурпурные бактерии (Rhodospirillales)
Общим для всех представителей Rhodospirillales является то, что их фотосинтетический аппарат (светособирающие системы и реакционные центры) находится на внутренних мембранах (тилакоидах), образующихся из впячиваний плазматической мембраны (см. рис. 2.23 и 2.24).
Таблица 12.1. Семейства анаэробных фототрофных бактерий
|
Порядок |
Семейство |
Типовой вид |
Рост |
|
|
аэробный в темноте |
анаэробный на свету |
|||
|
Rhodospirillales (пурпурные бактерии) |
Chromatiaceae (пурпурные серные бактерии) |
Chromatium vinosum |
_ 4 |
+ |
|
Rhodospirillaceae (пурпурные несерные бактерии) |
Rhodospirillum rubrum |
(+)5 |
+ |
|
|
Chlorobiales (зеленые бактерии) |
Chlorobiaceae (зеленые серные бактерии) |
Chlorobium limicola |
— |
+ |
|
Chloroflexaceae (зеленые несерные бактерии) |
Chloroflexus aurantiacus |
+ |
+ |
1 Бхл-бактериохлорофилл.
2 n-АБК-n-аминобензойная кислота.
3 ПМ-плазматическая мембрана.
4 Отдельные виды растут в темноте в аэробных или микроаэробных условиях. - Прим. ред.
5 Почти все виды растут в темноте в аэробных или микроаэробных условиях. - Прим. ред.
Морфология тилакоидных структур может сильно варьировать у разных видов. Различают везикулярные, трубчатые и пластинчатые формы (см. рис. 2.24). Типичным для данной группы бактерий хлорофиллом является (за немногими исключениями) бактериохлорофилл а. Все эти бактерии способны фиксировать СO2 в рибулозобисфосфатном цикле и использовать органические соединения как доноры водорода и (или) источники углерода.
По способности использовать в качестве донора электронов элементарную серу в группе пурпурных бактерий выделяют два семейства: пурпурные серные бактерии, или Chromatiaceae (прежнее название - Thiorhodaceae), и пурпурные несерные бактерии, или Rhodospirillaceae (прежнее название - Athiorhodaceae)1.
Пурпурные серные бактерии (Chromatiaceae). Большинство пурпурных серных бактерий (пурпурных серобактерий) можно легко распознать по внутриклеточным включениям серы в виде сильно преломляющих свет шариков (см. рис. 2.43, 12.2 и 12.3). Chromatium okenii (толщиной 5 мкм и длиной 20 мкм) и Thiospirillum jenense (толщиной 3,5 мкм и длиной 50 мкм) - гиганты среди бактерий (рис. 12.1, 12.2 и 12.3). Они давно уже привлекали к себе внимание микроскопистов; на них изучают движение с помощью жгутиков, а также двигательные реакции на различные раздражители. Chromatium warmingii отличается от С. okenii несколько меньшими размерами клеток и полярным расположением включений серы. Крупные представители этого рода имеют форму почки, а мелкие - коротких палочек. К ним относится и Chromatium vinosum, штамм D, на котором были проведены важные исследования по бактериальному фотосинтезу. Для Thiocystis (Т. violacea, Т. gelatinosa) характерны сферические подвижные клетки. У Thiocapsa roseopersicina и Т. pfennigii клетки тоже имеют сферическую форму, но они неподвижны. Многие виды пурпурных серобактерий обладают газовыми вакуолями. Таковы сферические подвижные бактерии Lamprocystis roseopersicina, а также неподвижные формы - Amoebobacter (сферические клетки), Thiopedia (эллипсоидные клетки), Thiodictyon (палочковидные клетки).
1 В настоящее время выделяют еще одно семейство - Ectothiorhodaceae; раньше соответствующие виды включали в семейство Chromatiaceae. - Прим. ред.
|
Xapaктерные признаки семейства |
||||
|
Окисление H2S |
Отложение серы |
Пигменты1,6 |
Фотосинтетический аппарат |
Потребность в витаминах |
|
+ |
Внутриклеточное |
Бхл а (Бхл b) |
Тилакоиды |
Нет или В12 |
|
-( + ) |
(Внеклеточное) |
Бхл а (Бхл b) |
Тилакоиды |
n-АБК2, тиамин, биотин, никотиновая кислота |
|
+ |
Внеклеточное |
Бхл а Бхл с Бхл d Бхл е |
ПМ3 Хлоросомы » » |
Нет или В12 |
|
( + ) |
Бхл а Бхл с |
ПМ3 Хлоросомы |
||
6 Недавно выделена анаэробная фототрофная бактерия Heliobacterium chlorum, осуществляющая аноксигенный фотосинтез. Она содержит особый хлорофилл (бактериохлорофилл д) и в отличие от других фототрофных бактерий является грам-положительной. Предполагают, что это наиболее древняя форма фототрофов. - Прим. ред.
Характерная особенность Chromatiaceae - то, что во время окисления H2S в их клетках в качестве промежуточного продукта откладывается сера. Представители Ectothiorhodospira (Е. mobilis, Е. halophila) откладывают серу не в цитоплазме, а вне клеток и там же окисляют ее далее до сульфата.
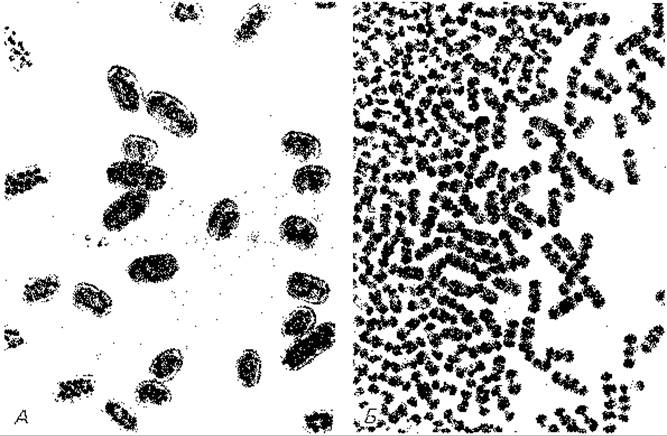
Рис. 12.2. Chromatium okenii (А) и Ch. warmingii (Б). Снимки в светлом поле; X ~ 800. (Фото N. Pfennig.)
Chromatiaceae обладают везикулярными тилакоидами (хроматофорами), которые обычно заполняют всю клетку. В настоящее время известны лишь два исключения: у видов Ectothiorhodospira тилакоиды образуют стопки ламелл, а у Thiocapsa pfennigii они имеют трубчатую форму.

Рис. 12.3. Thiospirillum jenense с монополярными политрихальными жгутиками и включениями серы. Снимок в светлом поле; X ~ 700. (Фото N. Pfennig.)
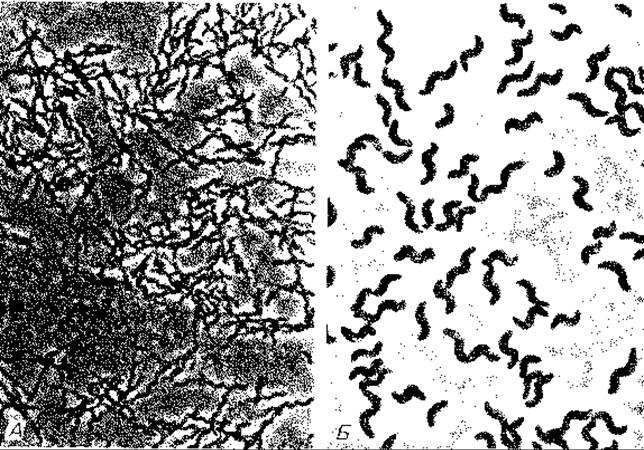
Рис. 12.4. Rhodospirillum rubrum (А) и Rh. fulvum (Б). Снимки в светлом поле; х ~ 1200 и ~ 1800 соответственно.
Пурпурные несерные бактерии (Rhodospirillaceae). Большинство известных бактерий этой группы можно отнести к двум родам. Палочкообразные виды объединяют в род Rhodopseudomonas; названия видов связаны главным образом с их физиологическими признаками: R. palustris, R. viridis, R. acidophila, R. blastica и R. sulfidophila. Спирилловидные формы относят к роду Rhodospirillum; по величине и окраске различают виды R. rubrum (рис. 12.4), R. salexigenes, R. fulvum, R. molischianum и R. photometricum. Тилакоидные структуры у пурпурных несерных бактерий весьма разнообразны по форме.
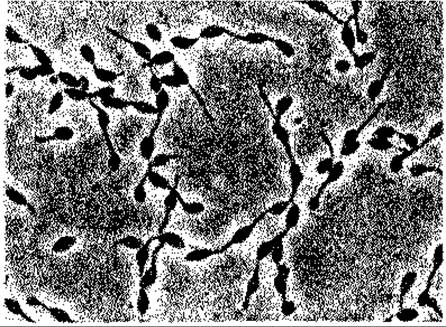
Рис. 12.5. Rhodomicrobium vannielii - несерная пурпурная бактерия, размножающаяся почкованием и образующая гифоподобные нити. Снимок в светлом поле; х ~ 1200. (Фото N. Pfennig.)
Особое положение среди Rhodospirillaceae занимают Rhodomicrobium vannielii и Rhodocyclus purpureus. Первый из этих видов (рис. 12.5) размножается почкованием; дочерние клетки остаются связанными с материнской клеткой при помощи гифоподобных нитей или же отделяются от нее, образуя клетки с перитрихально расположенными жгутиками. Rhodocyclus purpureus - единственная неподвижная форма в этом семействе; клетки имеют форму полукольца; аппарат фотосинтеза, вероятно, находится в плазматической мембране, у которой можно обнаружить лишь очень небольшие и малочисленные впячивания.
Сероводород подавляет рост многих пурпурных несерных бактерий; однако некоторые виды устойчивы к H2S или даже используют его в качестве донора водорода в процессе фиксации СЩ2. Rhodopseudomonas sulfidophila и R. palustris окисляют сероводород до сульфата, не образуя в качестве промежуточного продукта серу.
Зеленые бактерии (Chlorobiales)
Для представителей Chlorobiales характерно наличие хлоросом органелл, содержащих пигмент и прилегающих к плазматической мембране. В хлоросомах находится характерный для данной группы бактерий бактериохлорофилл (Бхл с, d или е) - светособирающий пигмент. Кроме того, эти бактерии имеют и небольшое количество бактериохлорофилла а, который прямо связан с фотосинтетическими реакционными центрами и локализован в плазматической мембране. Зеленые бактерии отличаются от пурпурных отсутствием рибулозобисфосфат-карбоксилазы; поэтому они не могут фиксировать СО2 в рибулозобисфосфатном цикле.
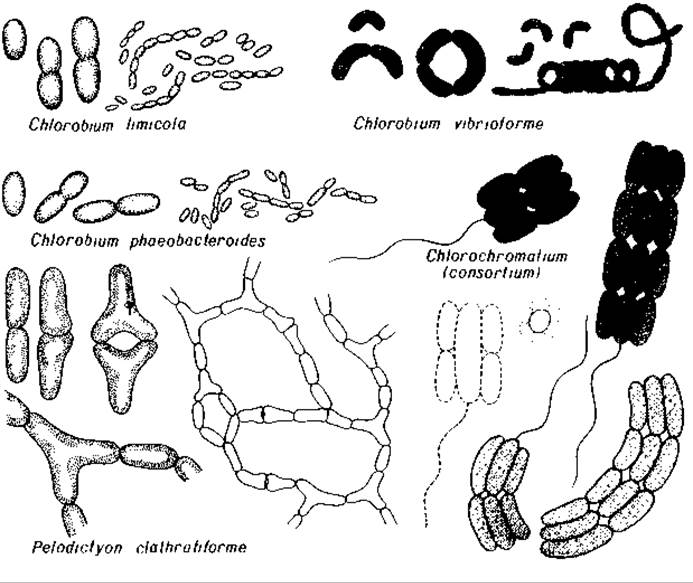
Рис. 12.6. Фототрофные зеленые серобактерии (Chlorobiaceae).
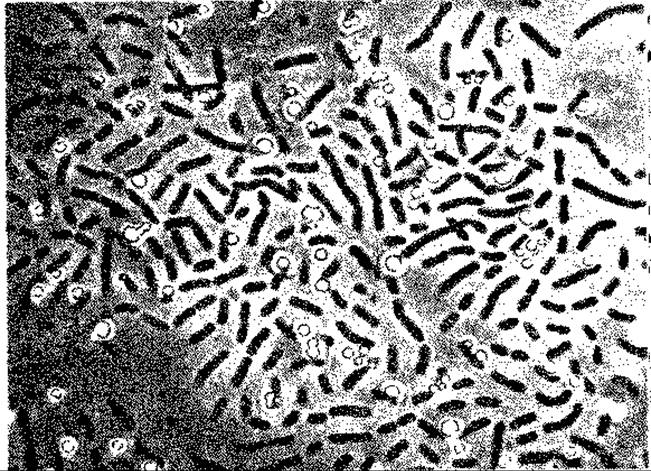
Рис. 12.7. Chłorobium limicola. Молодая культура с внеклеточными отложениями серы. Снимок в светлом поле; х ~ 1000. (Фото N. Pfennig.)
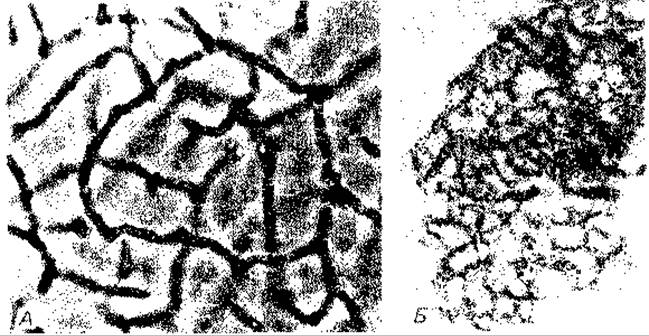
Рис. 12.8. Зеленая серная бактерия Pelndictyon clathratiforme (А), клетки которой соединены в сеть, и серная пурпурная бактерия Thiodictyon elegans (Б), образующая более рыхлую сеть. Снимки в светлом поле; х 1500 и 400 соответственно. (Фото N. Pfennig.)
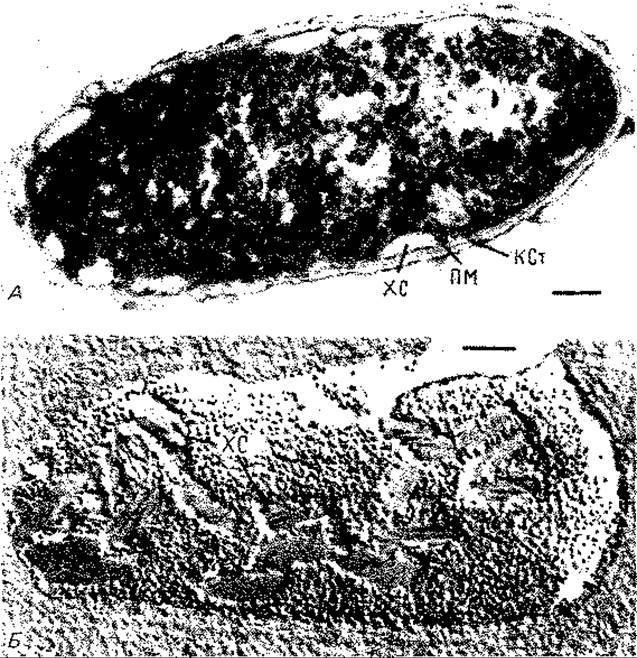
Рис. 12.9. Электронные микрофотографии Chlorobium limicola. А. Ультратонкий срез. Б. Хлоросомы, видимые благодаря технике замораживания-травления. ХС - хлоросомы; ЯМ - плазматическая мембрана; КСт - клеточная стенка. Длина балок около 100 нм. (Staehelin L. A. et al., Biochim. Biophys. Acta, 589 [1980], 30.)
К семейству Chlorobiaceae (рис. 12.6-12.9) относятся зеленые (Chlorobium vibrioforme и Ch. limicola) и окрашенные в коричневый цвет (Сh. phaeobacteroides) виды, а также формы, образующие звездчатые агрегаты (Prosthecochloris) или сети (Pelodictyon clathratiforme).
Chlorochromatium aggregatum представляет собой симбиотическую ассоциацию двух видов бактерий; бесцветных хемоорганотрофных палочек, передвигающихся с помощью длинных полярных жгутиков, и их эктосимбионтов - зеленых фототрофных бактерий. Сходную организацию имеет и Pelochromatium roseum с тем отличием, что эктосимбионт окрашен здесь в коричневый цвет. Морская бактерия Chloroherpeton thalassium отличается от видов Chlorobium нитевидной формой клеток и скользящим движением.
Chloroflexaceae. Фототрофная зеленая бактерия Chloroflexus по своей форме и способу передвижения относится к нитчатым скользящим бактериям, однако содержит бактериохлорофиллы с и а и обладает хлоросомами такого же типа, как у зеленых серобактерий (Chlorobiaceae). В то же время Chloroflexus отличается от видов Chlorobium своей способностью к аэробному гетеротрофному росту на сложных средах как на свету, так и в темноте. СЫогоПехасеае в отличие от Chlorobiaceae практически неспособны к фотоавтотрофному росту на СO2 и Н2, Таким образом, эти бактерии являются скорее фотогетеротрофами. С. aurantiacus-бактерия, широко распространенная во всем мире; это главный компонент зеленого или оранжевого мата на дне ручьев, берущих начало от горячих источников.
12.1.1 Пигменты фотосинтетического аппарата
Благодаря фотосинтетическим пигментам достаточно густые суспензии фототрофных бактерий имеют зеленую, сине-зеленую, пурпурно-фиолетовую, красную, коричневую или розовую окраску. Цвет зависит от природы и количественного соотношения пигментов. Отдельные пигменты можно распознать даже по спектрам поглощения интактных клеток (рис. 12.10). Хлорофиллы, например, ответственны за максимумы поглощения в синей (< 450 нм) и в красной и инфракрасной (650-1100 нм) областях спектра. Поглощение в области 400-550 нм обусловлено главным образом каротиноидами, а у цианобактерий в области 550-650 нм-фикобилипротеинами.
Различные хлорофиллы фототрофных микроорганизмов отличаются друг от друга в основном наличием или отсутствием двойной связи между углеродными атомами 3 и 4 и заместителями в порфириновом скелете (рис. 12.11). От этих модификаций зависят максимумы поглощения в ближней инфракрасной области. Различия отчетливо видны и в спектрах интактных клеток или изолированных фотосинтетических мембран, в которых пигменты образуют специфические комплексы с белками (рис. 12.10). Главный максимум хлорофилла а зеленых водорослей и цианобактерий лежит в области 680-685 нм, бактериохлорофиллов с, d и е зеленых серобактерий и Chloroflexus в области 715-755 нм, а бактериохлорофилла а большинства пурпурных бактерий - при 850-890 нм. В области от 1020 до 1035 нм поглощает бактериохлорофилл b, найденный пока лишь в клетках Rhodopseudomonas viridis, Ectothiorhodospira halochloris, Thiocapsa pfennigii и некоторых других пурпурных бактерий. Бактериохлорофилл а у пурпурных бактерий представлен четырьмя спектральными формами: В800, В820, В850 и В870-890. Различия в поглощении зависят в данном случае от характера связи и положения молекулы Бхл в пигментно-белковом комплексе фотосинтетического аппарата. Число пиков и их относительная высота изменяются от вида к виду, а у многих организмов на них влияют и условия культивирования.
Chlorobiaceae содержат наряду с характерными для них бактериохлорофиллами с, d или е также небольшое количество Бхл а лишь с одним пиком поглощения при 810 нм, тогда как в клетках Chloroflexus Бхл а дает два отчетливых пика при 808 и 868 нм.
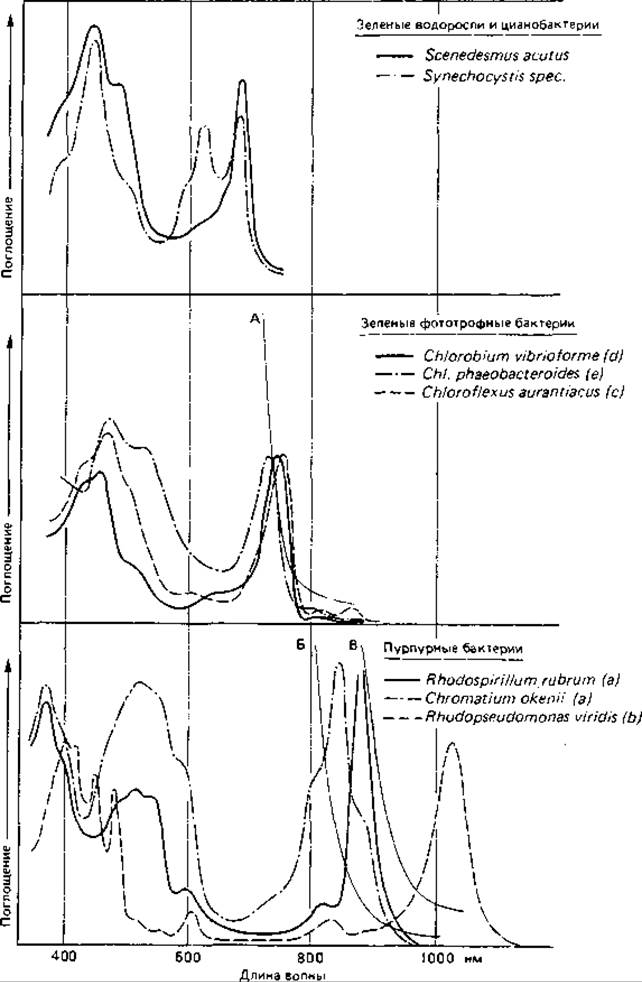
Рис. 12.10. Спектры поглощения интактных клеток фототрофных бактерий, цианобактерий и зеленых водорослей. В скобках указаны типы бактериохлорофиллов. Приведены также кривые, характеризующие способность к поглощению инфракрасных лучей у фильтров, используемых для получения элективных накопительных культур зеленых и пурпурных бактерий (А), пурпурных бактерий, содержащих бактериохлорофилл а (Б) и пурпурных бактерий, содержащих бактериохлорофилл b (В). (Pfennig N.. Ann. Rev. Microbiol. 21 [1967], 285.)
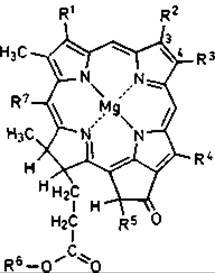
Рис. 12.11. Различия в структуре хлорофилла а и бактериохлорофиллов a, b, с, d и е.
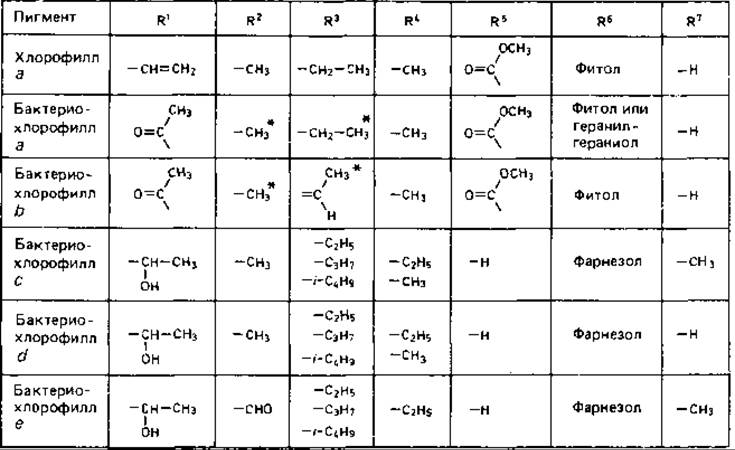
*Связь между С-3 и С-4 насыщенная
Каротиноиды являются так называемыми дополнительными пигментами фототрофных организмов. Они поглощают свет в спектральной области от 400 до 550 нм. У пурпурных бактерий это в основном алифатические С40-соединения (тегратерпеноиды) с третичными гидрокси- и метоксигруппами (ликопин, родопин, спириллоксантин, сфероидин). Оксогруппы и альдегидные группы могут придавать им темно-красную окраску (сфероиденон, окенон, родопиналь). Обнаружены также каротиноиды с ароматическими кольцами (арилкаротиноиды) - производные у- или ß-каротина. Эти каротиноиды встречаются у представителей лишь немногих родов (окенон - у некоторых Chromatiaceae; хлоробактин является типичным каротиноидом зеленых, а изорениера гин - коричневых видов Chlorobium).
Каротиноиды выполняют две функции: с одной стороны, они участвуют в фотосинтезе как светособирающие пигменты, т. е. поглощают световую энергию и передают ее хлорофиллу; с другой стороны, они предохраняют хлорофилл от фотоокисления. Сине-зеленые мутантные формы пурпурных бактерий, лишенные каротиноидов, способны расти только на слабом свету, а при высокой интенсивности света гибнут.
Из различий в спектрах поглощения зеленых водорослей, цианобактерий, пурпурных бактерий и зеленых бактерий можно заключить, что разные группы фототрофных организмов используют для фотосинтеза свет разных участков спектра. Это связано с условиями освещения в естественных местах обитания разных фототрофных организмов. Найденные различия используют при получении накопительных культур некоторых фототрофных бактерий (см. рис. 12.13).
Локализация пигментов. Фотосинтетические пигменты у пурпурных бактерий связаны с внутренними мембранами - везикулярными или трубчатыми выростами плазматической мембраны, которые сохраняют с ней связь, но проникают в толщу цитоплазмы. У разных видов бактерий такие мембраны имеют разную форму. Это могут быть трубочки, везикулы (пузырьки) или скопления ламелл (располагающихся концентрически или же в виде стопок); иногда они заполняют всю внутренность клетки (см. рис. 2.23). Фрагменты мембран, освобождаемые при разрушении клеток в виде везикул и отделяемые центрифугированием, называют «хроматофорами». В клетках зеленых бактерий пигменты связаны с различными структурами: светособирающие пигменты - главным образом с хлоросомами, а пигменты реакционных центров - с плазматической мембраной (см. рис. 2.4 и 12.9).
Регуляция синтеза пигментов и тилакоидов. Синтез фотопигментов зависит от условий роста организмов, прежде всего от освещенности и (у факультативных анаэробов) от присутствия кислорода. Содержание пигмента в клетках тем выше, чем ниже была освещенность во время их роста. Кислород тоже влияет на образование пигментов: так же как и яркий свет, он подавляет образование мембранных структур, содержащих пигменты, а тем самым и синтез бактериохлорофиллов и каротиноидов. Количества фотосинтетических пигментов изменяются параллельно с изменением числа везикул и трубочек (внутриклеточных мембран), выявляемых в клетках с помощью электронного микроскопа. Кислород, кроме того, ингибирует некоторые ферментативные этапы синтеза бактериохлорофилла. Наиболее высокое содержание фотопигментов, а также несущих пигмент везикул и трубочек можно обнаружить в клетках, выросших в анаэробных условиях при слабом освещении.
12.1.2 Метаболизм
Исследование обмена веществ у фототрофных бактерий сопряжено с рядом трудностей. Эти бактерии часто отличаются большой разносторонностью. Например, многие несерные пурпурные бактерии способны как к анаэробному росту на свету, так и к аэробному росту в темноте; другие группы представлены строгими анаэробами и облигатными фототрофами. Многие виды используют в качестве донора электронов молекулярный водород, а некоторые-сероводород или серу. Наблюдается ассимиляция СО2 и органических веществ. В анаэробных условиях в темноте за счет брожения может получаться небольшое количество энергии, недостаточное, однако, для роста большинства видов. Короче говоря, фототрофные бактерии относятся к организмам, наиболее разносторонним в отношении обмена веществ. Поэтому здесь мы можем лишь глобально охарактеризовать их физиологические возможности.
Фиксация СО2. Почти все до сих пор изученные фототрофные бактерии способны свызывать СО2 в рибулозобисфосфатном цикле. При этом для восстановления 3-фосфоглицерата пурпурные и зеленые бактерии используют не NADPH2 (как зеленые растения), a NADH2. Помимо этого в ассимиляции СО2 участвуют реакции восстановительного карбоксилирования, зависимые от ферредоксина или NAD(P). Фиксация СО2 либо обеспечивает возможность полностью автотрофного роста, либо при одновременном усвоении каких-либо сильно восстановленных органических соединений (например, жирных кислот) служит для поддержания определенного окислительно-восстановительного уровня веществ клетки. Виды Chlorobium ассимилируют СО2 через восстановительный цикл трикарбоновых кислот.
Доноры водорода. Анаэробные фототрофные бактерии нуждаются в экзогенном доноре водорода. Они способны использовать Н2, сероводород, элементарную серу, тиосульфат, органические кислоты, спирты, сахара и даже некоторые ароматические соединения. Молекулярный водород используется очень многими, но отнюдь не всеми фототрофными бактериями. Мелкие виды Chromatium, некоторые виды Rhodobacter (например, R. capsulata), родоспириллы и Chlorobium растут на свету, используя Н2 и СО2. Квантовый выход при этом примерно так же высок, как и при оксигенном фотосинтезе. Он составляет как у Rhodopseudomonas acidophila, так и у цианобактерии Anabaena cylindrica 8 молей квантов на 1 моль фиксированной двуокиси углерода.
Некоторые пурпурные несерные бактерии, пурпурные серные бактерии и зеленые серобактерии окисляют сероводород до сульфата. У большинства пурпурных серных бактерий в клетках в качестве промежуточного продукта временно откладывается сера. По-видимому, количественное преобладание крупных представителей Chromatium в естественных местообитаниях (прудах) связано с их способностью быстро окислять H2S на свету и аккумулировать серу; при этом внутриклеточная сера служит резервом восстановительной силы и позволяет ассимилировать СО2 на свету без поступления доноров водорода извне. В случае использования тиосульфата следует делать различие между сульфановой и сульфоновой серой; только сульфановая сера может откладываться в клетках, сульфоновая же поступает в окружающую среду в виде сульфата.
Некоторые виды Chlorobium в основном окисляют сероводород только до серы, которая выделяется в среду. Такие бактерии особенно хорошо растут в сообществе с Desulfuromonas acetoxidans: этот вид с «анаэробным дыханием» восстанавливает серу до H2S и окисляет при этом этанол или ацетат. Chlorobium и Desulfuromonas представляют типичный пример функциональной ассоциации, или синтрофии, двух микроорганизмов (см. разд. 9.3).
Темновой метаболизм. Многие пурпурные несерные бактерии и Сhlоroflexus способны к аэробному росту в темноте, если в их распоряжении имеются органические субстраты. Следовательно, они обладают компонентами дыхательного обмена, включая цикл трикарбоновых кислот. Последний участвует в метаболических превращениях также и в анаэробных условиях на свету. Использование большого числа органических кислот и сахаров некоторыми представителями Chromatiaceae и Rhodospirillaceae указывает на то, что метаболизм этих фототрофных бактерий хотя и варьирует, однако протекает в принципе по общеизвестным путям (таким, как фруктозобисфосфатный путь, путь Энтнера - Дудорова, цикл трикарбоновых кислот и др.).
Поскольку фототрофные бактерии должны выживать и ночью, не удивительно, что и в анаэробных условиях в темноте все же происходят энергетические процессы. Источником энергии является брожение, при котором донорами водорода служат резервные вещества, а акцептором водорода - сера (там, где она имеется). Конечные продукты анаэробного темпового метаболизма - СO2, ацетат, пропионат и сероводород. Однако в анаэробных условиях в темноте могут расти лишь отдельные виды фототрофных бактерий.
Образование Н2 на свету. Некоторые представители пурпурных бактерий способны на свету в присутствии подходящих органических или неорганических доноров водорода выделять молекулярный водород. Образование Н2 зависит от отношения С/N в субстратах, содержащихся в среде, и ингибируется свободными ионами аммония. N2тоже обратимо подавляет выделение клетками молекулярного водорода. Фотообразование Н2 связано с побочной функцией нитрогеназы; этот фермент способен восстанавливать наряду с N2также и протоны, что ведет к образованию молекулярного водорода. Поэтому при избытке энергии и восстановительной силы происходит выделение Н2.
Фиксация молекулярного азота. Подавляющее большинство исследованных пурпурных и зеленых бактерий способно фиксировать азот. Однако скорость их роста за счет использования N2, как правило, ниже, чем в присутствии ионов аммония.
Запасные вещества. Из запасных веществ для фототрофных бактерий характерны поли-β-гидроксимасляная кислота, полисахариды и полифосфаты. В зависимости от условий роста представители Chromatiaceae могут содержать также включения серы, которая находится в клетках в виде орторомбической серы.
12.1.3 Распространение фототрофных бактерий
Фототрофные пурпурные и зеленые бактерии обитают в анаэробных зонах многих водоемов, в мелководных прудах, медленно текущих водах, в озерах и морских бухтах. Пурпурные серобактерии нередко образуют налеты, окрашенные во всевозможные оттенки красного цвета, от нежно-розового до темно-красного, на поверхности ила или на каком-нибудь разлагающемся растительном материале. Иногда они роятся над поверхностью ила, образуя слой толщиной около дециметра. Такое «цветение» воды в мелких водоемах (рис. 12.12) вызывают в первую очередь крупные представители пурпурных бактерий - Chromatium okenii С. warmingii, С. weissei и Thiospirillum jenense, однако встречаются и мелкие представители Chromatiaceae и Chlorobiaceae. Бурное размножение пурпурных серобактерий наблюдается и в таких мелководных прудах, поверхность которых покрыта плотным слоем ряски (Lemna) или же листьями водяных лилий. Этот своеобразный биологический фильтр поглощает те спектральные компоненты света, которые могли бы использоваться зелеными водорослями и цианобактериями, но пропускает свет, поглощаемый бактериохлорофиллами и темно-красными каротиноидами. Поэтому под покровом из ряски растут анаэробные фототрофные бактерии, но не развиваются зеленые водоросли и цианобактерии (рис. 12.12). Некоторые виды фототрофных серобактерий представлены в естественных местообитаниях почти чистой культурой. Всегда можно обнаружить и Rhodospirillaceae, но лишь редко в столь массовом количестве.
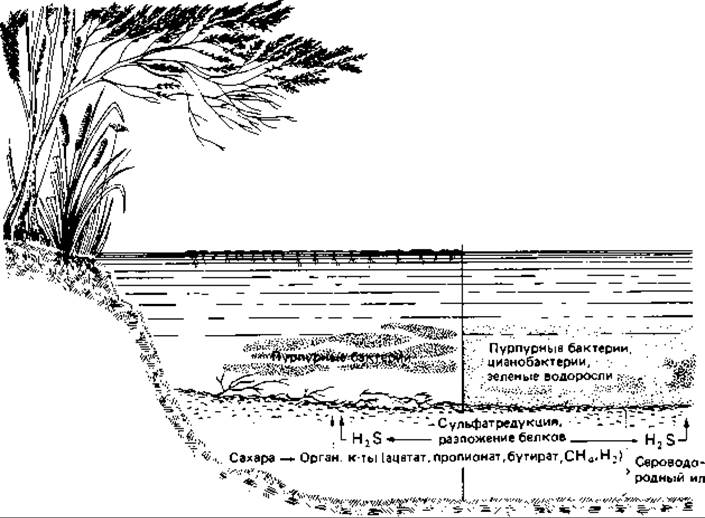
Рнс. 12.12. Профиль неглубокого водоема, где наблюдается массовое развитие серных пурпурных бактерий.
Сезонное массовое развитие пурпурных серных бактерий наблюдается также в анаэробных зонах озер ниже области температурного скачка (термоклина, см. рис. 17.1). Бактерии находят здесь необходимые им вещества - сероводород, СO2 и органические соединения. Инфракрасное солнечное излучение, конечно, не проникает на глубину 10-30 м; на такой глубине максимум энергии приходится на синюю и сине-зеленую области спектра (450-500 нм), т. е. именно на ту область, где поглощают каротиноиды. Это делает понятным относительно высокое содержание каротиноидов у пурпурных бактерий, определяющее их окраску: каротиноиды (подобно фикоэритринам у красных водорослей и цианобактерий) делают возможным у пурпурных бактерий фотосинтетический метаболизм на больших глубинах. Соответственно на таких глубинах и среди зеленых серобактерий преобладают богатые каротиноидами коричневые формы (Chlorobium phaeobacteroides, С. phaeovibrioides, Pelochromatium).
12.1.4 Накопительные культуры
Метод отбора анаэробных фототрофных бактерий основан на том (описанном выше) наблюдении, что пурпурные серные бактерии достигают иногда массового развития и в мелких водоемах, если вся поверхность воды покрыта плотным слоем ряски. С помощью фильтров, поглощающих коротковолновую часть спектра и пропускающих инфракрасные лучи, используемые лишь отдельными группами фототрофных бактерий, можно создать селективные условия для роста как зеленых серобактерий, так и пурпурных бактерий, содержащих бактериохлорофилл а или b.
Будут ли в накопительной культуре («колонке Виноградского», рис. 12.13) развиваться преимущественно пурпурные или зеленые бактерии, зависит от имеющихся доноров водорода и от концентрации H2S. Если цилиндр, в котором находятся песок, почва и яичный белок, заполнить водой, а затем внести в него инокулят из места обитания фототрофных бактерий, то на свету будут развиваться виды Rhodospirillaceae. Если же, добавив сульфат кальция, обеспечить постоянное образование H2S в результате восстановления сульфата, то рост пурпурных несерных бактерий будет подавлен и доминировать будут серные бактерии. В синтетических питательных растворах, содержащих витамин В12, можно получать накопительные культуры различных видов зеленых и пурпурных бактерий, более тонко варьируя такие факторы, как концентрация сероводорода и питательных солей, pH, температура и интенсивность света. Формы, отличающиеся особой чувствительностью к условиям среды, например бактерии, образующие газовые вакуоли, нуждаются обычно в более низких концентрациях H2S, более низких температурах и меньшей интенсивности света. При получении накопительных культур отдельных видов Rhodospirillaceae решающую роль играют выбор донора водорода и присутствие некоторых витаминов (биотина, и-аминобензойной кислоты, тиамина, никотиновой кислоты).
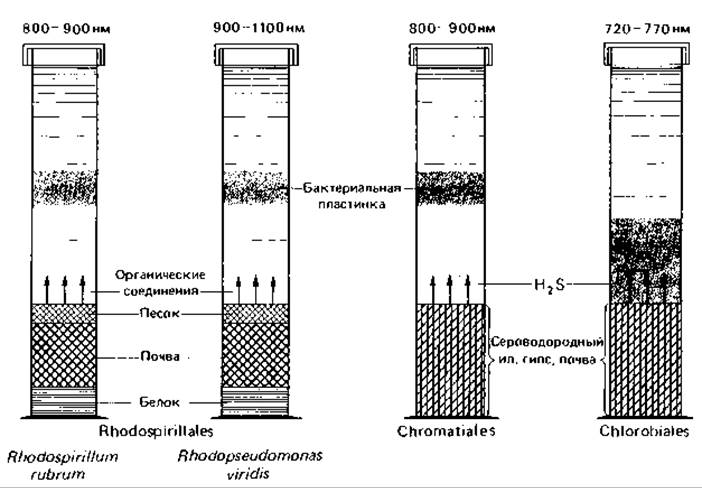
Рис. 12.13. Накопительные культуры фототрофных бактерий в «колонках Виноградского». Числа над колонками отражают спектральные характеристики светофильтров, под которыми наблюдается предпочтительное развитие указанных видов и групп бактерий. Через несколько дней после заполнения колонок и засева среды прудовым илом и водой в толще воды образуются красные или зеленые «бактериальные пластинки».