ЗАГАЛЬНА ГІСТОЛОГІЯ З ОСНОВАМИ ЕМБРІОЛОГІЇ - Долгов О. М. - 2015
Частина І
БІОЛОГІЯ КЛІТИНИ
ЯДРО
Ядерце
Практично в усіх живих клітинах еукаріотичних організмів в ядрі виявляється одне або декілька зазвичай кулястої форми тілець розмірами 1-5 мкм, які сильно заломлюють світло і добре профарбовуються лужними барвниками. Така базофілія зумовлена тим, що ядерця багаті на РНК. Незалежно від рівня диференціювання, чи в недиференційованих клітинах ембріональних, тих, що регенерують, або пухлинних тканин, чи в диференційованих клітинах, що синтезують велику кількість білка (наприклад, клітини залозистого епітелію, нейрони, плазмоцити і т.п.), цитоплазма яких багата на РНК, виявляються великі базофільні ядерця.
Разом з тим, ядерце не є самостійною структурою або органоїдом. Ядерце є похідним хромосоми, одним з її локусів, що активно функціонує в інтерфазі.
Утворення ядерець та їх кількість пов’язані з активністю та численністю певних ділянок хромосом - ядерцевих організаторів, зміни кількості ядерець у клітинах даного типу можуть відбуватись за рахунок злиття ядерець або за рахунок зміни числа хромосом з ядерцевими організаторами. ДНК ядерцевого організатора представлена поліцистронним геном рРНК (декілька сотень копій). Подібне явище багаторазового повторювання одного гена з метою мультиплікації синтезу певної молекулярної субстанції визначається терміном ампліфікація. На кожному з цих генів синтезується високомолекулярна рРНК-попередник, яка потім перетворюються у коротші молекули рРНК, що входять до складу субодиниць рибосом (Рис. 20).
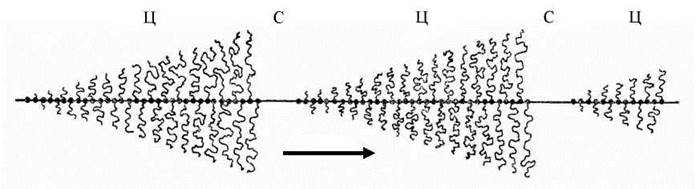
Рис. 20. Схема роботи ядерцевого організатора.
Ц - цистрони, С - проміжки, на яких відсутній синтез рРНК. Стрілка показує напрямок, в якому вздовж молекули ДНК рухається молекула РНК-полімерази І (чорні крапки на молекулі ДНК), від якої відходить синтезована молекула рРНК (звивиста лінія).
Схему участі ядерець у синтезі цитоплазматичних білків можна уявити собі наступним чином (Рис. 21): спочатку на ДНК-матриці за допомогою РНК-полімерази І синтезується 45S-попередник рРНК, потім 45Б-попередник рРНК взаємодіє з рибосомними білками з наступним розділенням на 28S, 18S і 5,8S рРНК, рибонуклеопротеїни, які містять 28S і 5,8S рРНК, об’єднуються з 5S рРНК, що синтезується поза ядерцем, і утворюють велику субодиницю рибосоми (СО), рибонуклеопротеїни, які містять 18S рРНК, формують малу субодиницю рибосоми.
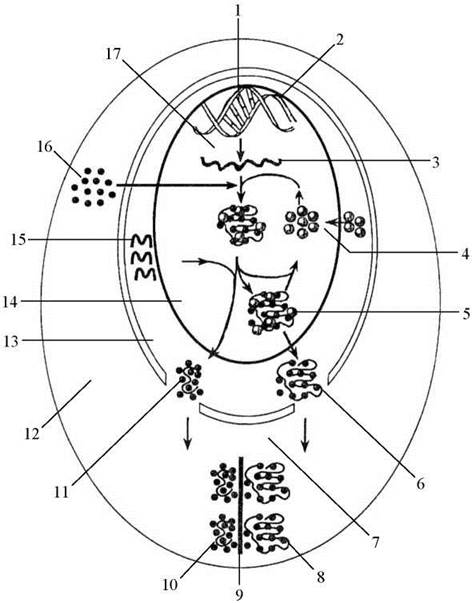
Рис. 21. Участь ядерця в утворенні рибосом.
1 - ДНК, 2 - петля ядерцевого організатора, 3 — 45S-попередник рРНК, 4 - колообіг РНК і білків, утягнених в процесинг, 5 - незріла велика СО, 6 - велика СО, 7 - транспорт і активація, 8 — 60S-субодиниця, 9 - мРНК, 10 — 40S-субодиниця, 11 - мала СО, 12 - цитоплазма, 13 - ядро, 14 - ядерце, 15 - 5S рРНК, синтезована поза ядерцем, 16 - рибосомні білки, що утворюються в цитоплазмі, 17 - транскрипція.
ДНК ядерцевого організатора з синтезованими на її матриці молекулами рРНК, вкрита ззовні гранулами РНП, що перетворюються на СО рибосом, має товщину близько 200 нм і називається нуклеонемою. В її складі гранули мають розміри близько 15-20 нм, а фібрилярний компонент - нитки рРНК - 6-8 нм. Нуклеонема скручена в клубочок кулястої форми. Через те, що матриця ДНК, вкрита ззовні фібрилярним і гранулярним компонентами, якісна гістохімічна реакція на ДНК (реакція Фельгена) в зоні ядерця дає негативний результат.
На поперечному розрізі нуклеонеми електронна мікроскопія виявляє три її структурних елементи: фібрилярний центр - слабозабарвлений компонент, представлений ДНК, що кодує рРНК; фібрилярний компонент, де відбуваються ранні стадії утворення попередників рРНК (складається з тонких діаметром 5 нм рибонуклеопротеїнових фібрил і транскрипційно активних ділянок ДНК); гранулярний компонент містить зрілі попередники СО рибосом діаметром 15 нм.
Транскрипція рРНК в ядрах клітин людини відбувається в хромосомах 13, 14, 15, 21 та 22. Петлі ДНК саме цих хромосом, які містять відповідні гени, і формують ядерцевий організатор.