ЗАГАЛЬНА ГІСТОЛОГІЯ З ОСНОВАМИ ЕМБРІОЛОГІЇ - Долгов О. М. - 2015
Частина І
БІОЛОГІЯ КЛІТИНИ
ЯДРО
Ядерна оболонка
Ця структура характерна для всіх еукаріотичних клітин. Ядерна оболонка складається з зовнішньої і внутрішньої мембран, розділених перинуклеарним простором (люмен).
Мембрани ядерної оболонки в морфологічному відношенні не відрізняються від інших внутрішньоклітинних мембран: вони мають звичайну ліпопротеїнову будову і завтовшки близько 7 нм. Обидві ядерні мембрани розділені перинуклеарним простором завширшки 20-60 нм.
У загальному вигляді ядерна оболонка може бути представлена як порожній двошаровий мішок, що відокремлює вміст ядра від цитоплазми. З усіх внутрішньоклітинних мембранних компонентів таким типом розташування мембран володіють тільки ядро, мітохондрії і пластиди. Однак ядерна оболонка має характерну рису, що відрізняє її від інших мембранних структур клітини. Це наявність особливих пор в оболонці ядра, що утворюються за рахунок численних зон злиття двох ядерних мембран і являють собою як би округлі перфорації всієї ядерної оболонки.
Зовнішня мембрана ядерної оболонки, що безпосередньо контактує з цитоплазмою клітини, має ряд структурних особливостей, що дозволяють віднести її до власне мембранної системи ендоплазматичного ретикулума. Так, на зовнішній ядерній мембрані звичайно розташовується велика кількість рибосом, як і на мембранах ергастоплазми. Існують численні спостереження про безпосередній перехід зовнішньої ядерної мембрани в систему каналів ергастоплазми, що особливо підкреслює структурну ідентичність цих мембран. У більшості тваринних і рослинних клітин зовнішня мембрана ядерної оболонки не являє собою ідеально рівну поверхню - вона може утворювати різної величини випинання або вирости убік цитоплазми. Такі вирости можуть мати вигляд пухирців або довгих трубчастих утворень.
Внутрішня мембрана контактує з хромосомним матеріалом ядра. Майже в усіх клітинах безпосередньо під внутрішньою ядерною мембраною розташований фіброзний, або щільний, шар - ядерна пластинка, або ламіна, завтовшки 80-300 нм. Ламіна містить білки проміжних філаментів - ламіни А, В і С.
У профазі три білки ядерної ламіни сильно фосфорилюються. Саме це, як думають, приводить до розпаду ламіни. Скоріше за все, у результаті цього розпадається і вся ядерна мембрана. Невеликі замкнуті пухирці, що утворюються з неї, морфологічно не відрізняються від елементів ендоплазматичного ретикулуму, їх добре видно під час мітозу навколо мітотичного апарата. Можливо, що частина комплексів ядерних пор під час мітозу залишається зв’язаною з хромосомами, але це вірогідно не встановлено.
У телофазі фрагменти ядерної мембрани зв’язуються з поверхнею окремих хромосом, частково оточують кожну з них і тільки після цього зливаються, утворюючи повну ядерну мембрану, одночасно відновлюються ядерні пори, а дефосфорильовані білки ламіни знову агрегують з утворенням інтактної структури. Можливо, що процес відновлення ядерної мембрани регулюється реполімеризацією білків ламіни, тим більше, що один з них після профази залишається зв’язаним із фрагментами ядерної мембрани і може, таким чином, мітити ці фрагменти для використання при побудові дочірніх ядер.
Ламіна взаємодіє не тільки з внутрішньою ядерною мембраною, але також із хроматином і з ядерними порами, тому цикл фосфорилювання-дефосфорилювання білків ламіни під час мітозу цілком міг би ініціювати відповідно розпад старої й утворення нової ядерної мембрани.
Найбільш характерною і структурою, що кидається в очі, у складі ядерної оболонки є ядерна пора. Пори в оболонці утворюються за рахунок злиття двох ядерних мембран у вигляді округлих наскрізних отворів або перфорацій.
Ядерна оболонка клітин ссавців містить 3-4 тисячі пор (приблизно 10 пор на 1 квадратний мкм), організованих за участю спеціалізованих білків - нуклеопоринів. Через ядерні пори відбувається обмін речовинами між ядром і цитоплазмою. Дійсно, РНК, синтезовані в ядрі, а також рибосомні субодиниці і білки, що містять сигнали ядерного експорту, транспортуються через ядерні пори в цитоплазму, а гістони, компоненти реплікативної системи, багато інших білків і малі ядерні РНП імпортуються через ядерні пори з цитоплазми в ядро.
Пори оточені великими кільцевими структурами, називаними комплексами пори (їхній внутрішній діаметр складає приблизно 80 нм, а молекулярна маса - 50-100 мД). Кожен комплекс утворений набором великих білкових гранул, згрупованих в октагональну структуру (Рис. 22).
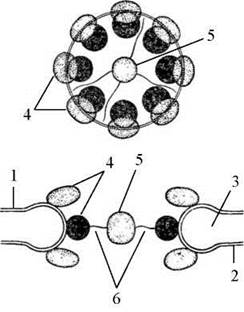
Рис. 22. Організація ядерної пори.
1 - зовнішня ядерна мембрана, 2 - внутрішня ядерна мембрана, 3 - люмен, 4 - периферичні гранули, 5 - центральна гранула, 6 - діафрагма пори.
Комплекс пори пронизує подвійну мембрану, зв’язуючи по окружності пори ліпідний бішар внутрішньої і зовнішньої мембран у єдине ціле. Незважаючи на цю безперервність, що повинна була б забезпечувати дифузію компонентів між зовнішньою і внутрішньою мембранами, вони залишаються хімічно різними.
Отвір у центрі кожного комплексу (ядерна пора) являє собою водний канал, крізь який водорозчинні молекули курсують між ядром і цитоплазмою.
Ядерний комплекс пори містить заповнений водою циліндричний канал діаметром близько 9 нм. Великі ядерні білки взаємодіють з білками-рецепторами, розташованими на границі ядерних пор, і ці рецептори активно переносять білки в ядро, збільшуючи канал пори.
Ядерні пори - утворення масою від порядку 66 мД в дріжджів до 125 мД у вищих еукаріот. Кількість білків (нуклеопоринів) у складі ядерної пори також варіює для різних організмів від 30 (приблизно) у дріжджів, до 50-100 видів білків у вищих еукаріот. На одне ядро припадає близько 190 ядерних пор у дріжджів, 3000-5000 у клітинах людини, що поділяються, і порядку 50 мільйонів у зрілих ооцитах Xenopus (шпорцева жаба). Утім, кількість ядерних пор залежить від типу клітини, стадії клітинного циклу і конкретної гормональної ситуації. Для ядерної пори характерна симетрія восьмого порядку, тому багато білків ядерної пори представлені в її складі в кількості, кратному восьми.
Лінійні розміри ядерних пор з різних об’єктів зазначені на Рис. 23.
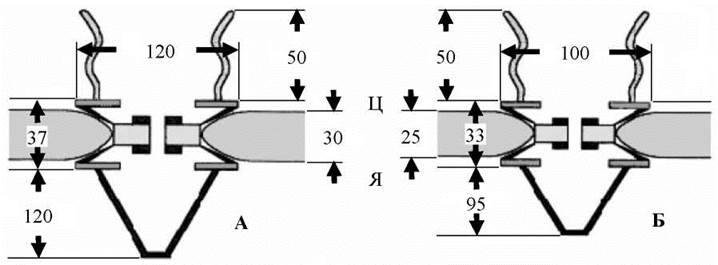
Рис. 23. Лінійні розміри ядерних пор, нм: А - шпорцевої жаби, Б - дріжджів.
Ц - цитоплазматична поверхня, Я - ядерна поверхня ядерної оболонки.
І на цитоплазматичній, і на ядерній стороні пори в електронний мікроскоп помітні опуклі кільця. Кільце, що знаходиться з ядерного боку, несе структуру, називану кошиком (basket). Це утворення складається зі звернених у нуклеоплазму фібрил і прикріпленого до них термінального кільця. До цитоплазматичного кільця також прикріплені фібрилярні білки. До просвіту каналу звернені вісім симетричних утворень (spoke complex), що додають комплексові подібності з восьмиспицевим колесом воза. Домени цих восьми субодиниць, що знаходяться в люмені, очевидно, зв’язуються між собою, закріплюючи структуру. У центрі комплексу видно вхід у канал ядерної пори. Іноді в каналі виявляється електроннощільна гранула. Деякі дослідники думають, що це якийсь комплекс, що транспортується, у момент перетинання ядерної мембрани. Інші вважають, що ця структура є функціональною деталлю ядерної пори. На підставі цього останнього припущення була навіть висунута гіпотеза, що згодом не підтвердилася, відповідно до якої ядерна пора містить не один, а вісім проникних каналів (Fahrenkrog, B. et. al. 1998 і T. Danker, H. Oberleithner, 2000).
Як уже згадувалося, ядерна пора проникна для дифузії. Молекули масою менше 5 кД проходять через ядерну пору вільно, і рівновага між ядерною і цитоплазматичною концентрацією встановлюється за секунди. Для білків масою 17 кД цей процес займає 2 хвилини, білків масою 44 кД (приблизно 6 нм у діаметрі) - 30 хвилин. Білки масою більш 60 кД, очевидно, взагалі не можуть пасивно проходити через ядерні пори. Проникний для гідрофільних макромолекул канал, через який відбувається як активний, так і пасивний транспорт, у ядерній порі один, і він, як видно, розташований у центрі комплексу. Параметри центрального каналу залежать від методів визначення. Результати, отримані різними методами і на різних об’єктах, коливаються в межах від 6,8 до 13 нм для діаметра еквівалентного каналу, і від 15 до 63,5 нм для його довжини. Під еквівалентним каналом розуміється циліндричний стовп води, аналогічний каналові ядерної пори по проникності для речовин, які пасивно дифундують; тобто ці чисельні розрахунки робилися без урахування властивостей білкових стінок каналу. З огляду на радіус каналу, абсолютно неясно, яким чином проходять через ядерні пори субодиниці рибосом масою 1,4 і 2,8 мД. З іншого боку, виявлено, що через ядерні пори можуть проходити частинки колоїдного золота діаметром до 26 нм.
Методами електронно-мікроскопічного аналізу показано, що ядерна пора досить лабільна структура. У відповідь на багато стимулів вона може змінювати свій радіус і, можливо, провідність. Було виявлено, що підвищення концентрації кальцію і ATФ можуть приводити до зменшення радіуса ядерної пори і збільшенню її висоти над рівнем мембрани. Оскільки просвіт (люмен) двомембранної оболонки ядра безупинно переходить у багатий на кальцій ендоплазматичний ретикулум, існує припущення, що вихід іонів Ca2+ у цитоплазму може грати безпосередню регуляторну роль (T. Danker, H. Oberleithner, 2000). Ця досить несподівана теорія якоюсь мірою доповнюється даними про перебування в складі ядерної пори скоротливого білка міозину, функція якого залежить від кальцію і ATФ. У ході активної транслокації комплекс вантаж-транспортини якориться на білках ядерної пори.
Процес транслокації субстрату через ядерну пору (для випадку імпорту) був досліджений методами електронної мікроскопії з застосуванням часток колоїдного золота (Рис. 24). На підставі цих даних процес можна розбити на кілька стадій. На першій стадії комплекс, що транспортується, якориться на зверненій у цитоплазму фібрилі (1). Потім цей філамент згинається і переміщує комплекс до входу в канал ядерної пори (2-3). Відбувається власне транслокація і звільнення комплексу в нуклеоплазму (4) (Pante N., Aebi U. 1996).
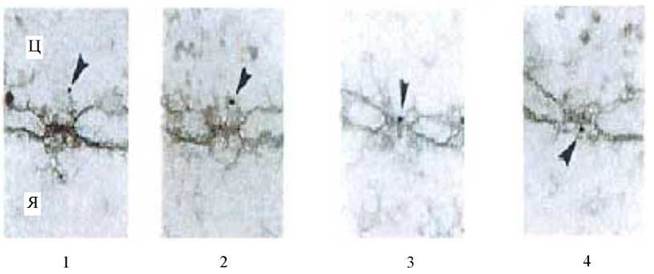
Рис. 24. Транслокація (імпорт) частинки колоїдного золота через ядерну пору.
Ц - цитоплазма, Я - ядро. Стрілкою показана частинка колоїдного золота.
Пояснення в тексті.