Введение в молекулярную биологию: От клеток к атомам - Энтони Рис, Майкл Стернберг 2002
Нуклеиновые кислоты и гены
Генетический код
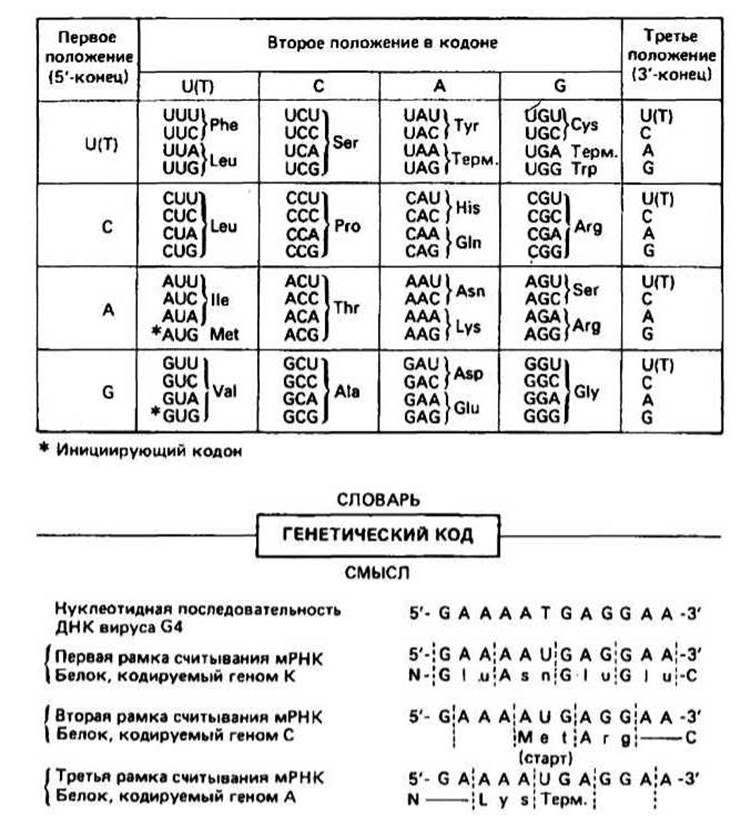
Рис. 23.1.
Генетический код связывает последовательность оснований данного гена или его РНК-транскрипта с аминокислотной последовательностью соответствующего белка.
Код триплетный, т. е. одна аминокислота задается последовательностью из трех нуклеотидов, называемой кодоном. В ДНК имеются четыре основания, а в белках — 20 аминокислотных остатков; синглетный код мог бы кодировать только четыре аминокислотных остатка, дублетный 4 ∙ 4 = 16 аминокислот, а триплетный образует 4 ∙ 4 ∙ 4 = 64 разных кодона.
Код не перекрывается т. е. в последовательности оснований ABCDEFGHI первые три основания, ABC, кодируют аминокислоту 1, DEF — аминокислоту 2 и т. д. Если бы код был перекрывающимся, то последовательность ABC кодировала бы аминокислоту 1, CDE - аминокислоту 2 и т. д. Неперекрывающийся характер кода относится только к случаю, когда рамка считывания не меняется (см. ниже). В коде отсутствуют запятые, т. е. нет знаков, отделяющих один кодон от другого.
Направление чтения закодированной записи — от 5'-конца к 3'-концу мРНК, являющейся транскриптом «+»-цепи ДНК, считанным с нее в направлении 5' -*3'. Первый с 5'-конца кодон отвечает N-концевой аминокислоте полипептидной цепи. Следовательно, белки синтезируются от N-конца к С-концу. Таблица кода указывает, какая аминокислота кодируется тем или иным кодоном (рис. 23.1). В таблице на рис. 23.1 первый (с 5'-конца) нуклеотид помещен в левом столбце, второй — в верхней строке, а третий (на 3'-конце) — в правом столбце. Из представленных в таблице 64 кодонов 61 кодон детерминирует ту или иную аминокислоту (например, CUA отвечает лейцину Leu), а три остальных кодона являются сигналами терминации (Терм.). Эти три кодона называются нонсенс-кодонами, поскольку они не определяют никакой аминокислоты. Помимо этого, когда кодоны
AUG для Met и иногда GUG для Val находятся в начале последовательности и соответствующие аминокислоты должны быть помещены в начале белковой цепи, эти аминокислоты обычно присутствуют в виде N-формильных производных (гл. 25). Когда же эти кодоны находятся в любом другом месте последовательности, то в пептидную цепь включаются нормальные Met и Val.
Таблица 23.1
|
Число разных кодонов |
Аминокислотные остатки |
|
6 |
Arg, Leu, Ser |
|
4 |
Ala, Gly, Pro, Thr, Val |
|
3 |
He (Терм.) |
|
2 |
Asn, Asp, Cys, Gin, Glu, His, Lys, Phe, Tyr |
|
1 |
Met, Trp |
Код вырожден, т. е. большинство аминокислот кодируется более чем одним кодоном (см. табл. 23.1). Например, Phe кодируется двумя кодонами, UUU и UUC. Кодоны, которые определяют одну и ту же аминокислоту, называются кодонами-синонимами. Вырожденность кода, как правило, выражается в том, что у кодонов, определяющих одну и ту же аминокислоту, первые два основания фиксированы, а третье положение может занимать одно из двух, трех или четырех разных оснований. В частности, кодоны с одним из двух пиримидинов (С или U) в третьем положении всегда являются синонимами, в то время как кодоны с одним из двух пуринов (А и G) в третьем положении бывают синонимами лишь иногда. Различия по всем трем положениям наблюдаются лишь в некоторых случаях (например, UCG и AGU оба кодируют Ser).
Гипотеза «качания» была предложена Криком для объяснения вырожденности кода по третьему положению основания в кодоне. Эта гипотеза, впоследствии получившая подтверждение, состоит в том, что соответствие третьего основания кодона первому основанию антикодона тРНК является нестрогим. Часто первое положение в антикодоне тРНК бывает занято необычным основанием инозином (I), которое может образовывать водородные связи с U, С или А, находящимися в кодоне в третьем положении (рис. 23.2). Полный набор образующихся при этом пар приведен в табл. 23.2. В связи с механизмом «качания» клетке требуется меньше 64 разных тРНК. Каждая тРНК может узнавать до трех кодонов.
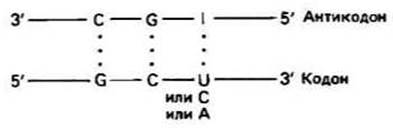
Рис. 23.2.
Таблица 23.2
|
Первое основание антикодона Третье основание кодона |
|
|
I |
U, С или А |
|
U |
А или G |
|
G |
С или U |
Химические свойства разных аминокислот находят отражение в структуре кода. Все кодоны с U во втором положении кодируют аминокислоты с гидрофобной боковой цепью (Phe, Leu, He, Met и Val). Если исключить терминирующие кодоны, то наличие А во втором положении определяет полярную или заряженную боковую цепь (Туг, His, Gin, Asn, Lys, Asp и Glu).
Рамка считывания задает положение первого основания кодона мРНК (или гена). Поскольку код три-плетен, число возможных рамок считывания равно трем. Обычно функциональный белок синтезируется только при одной рамке считывания, но некоторые вирусы используют две или даже три рамки считывания, при этом синтезируются разные белки (гл. 27). Примером такого рода могут служить белки, кодируемые К-, С- и А-генами вируса G4 (рис. 23.1).
Мутация — это изменение в последовательности оснований генетического материала данного организма. Знание генетического кода позволяет объяснить эффект некоторых мутаций.
Молчащая мутация — это такое изменение в нуклеотидной последовательности, которое приводит к образованию синонимичного кодона, и в результате аминокислотная последовательность кодируемого белка не изменяется. Структура кода такова, что молчащие мутации часто бывают обусловлены изменениями оснований лишь в третьем положении кодона.
Замена (миссенс-мутация) ведет к замещению одной аминокислоты другой в результате такого изменения последовательности оснований, которое не приводит к образованию синонимичного кодона. Так, заболевание серповидноклеточная анемия (гл. 16) возникает в результате замены Glu на Val в шестом положении (3-цепи гемоглобина человека. Это обусловлено изменением кодона GAA на GUA, т. е. заменой А на U во втором положении.
Мутация со сдвигом рамки обусловлена вставкой или удалением (делецией) одного или большего числа оснований в последовательности, так что при этом изменяется рамка считывания. Это приводит к изменению аминокислотной последовательности белка от точки мутации до С-конца молекулы.
Универсальность генетического кода означает, что все живые организмы — эукариоты, прокариоты и вирусы - используют один и тот же код. Хотя, вообще говоря, это положение справедливо, проведенные сравнительно недавно (1981 г.) определения нуклеотидной последовательности митохондриальных ДНК человека и дрожжей выявили некоторые необычные факты. Например, триплет UGA не является терминирующим кодоном, а кодирует Тгр, а триплеты AGA и AGC не кодируют Arg, а являются терминаторами. Генетический код был расшифрован в начале 60-х годов Ниренбергом, Кораной и их сотрудниками. Ниренберг получил клеточный экстракт Е. сой, содержащий все компоненты, необходимые для синтеза белка, включая рибосомы, все тРНК и аминоацил-тРНК-синтетазы (гл. 24). В систему добавляли полинуклеотид poly(U) (т. е. UUUUUUU...), функционировавший как искусственная мРНК. Оказалось, что poly(U) детерминирует синтез poly(Phe) (т. е. Phe, Phe, Phe...); следовательно, триплет UUU кодирует Phe. Аналогичные опыты показали, что триплет ССС кодирует Pro, а триплет ААА — Lys. Следующий шаг заключался в использовании полирибонуклеотидов, содержавших два, три или четыре разных основания, расположенных в случайном порядке. В результате этих исследований удалось определить состав других кодонов, но не последовательность оснований в них; так, было показано, что кодон, содержащий 2U и 1G, детерминирует Cys, но порядок оснований оставался неизвестным. Корана использовал полирибонуклеотиды не со случайной, а с заранее заданной последовательностью и определил структуру нескольких кодонов. Эти исследования получили дальнейшее развитие с помощью другого подхода, разработанного Ниренбергом. Он обнаружил, что тринуклеотиды вызывают связывание специфичных аминоацил-тРНК с рибосомой (гл. 24). Например, в присутствии UGU с рибосомой связывается только аминоацил-тРНК для Cys. Следовательно, UGU кодирует Cys. Три кодона, UAA, UAG и UGA, не кодируют никаких аминокислот и называются нонсенс-кодонами. Позднее исследования на мутантных бактериофагах показали, что эти кодоны определяют терминацию синтеза.