Введение в молекулярную биологию: От клеток к атомам - Энтони Рис, Майкл Стернберг 2002
Нуклеиновые кислоты и гены
Упаковка генетического материала
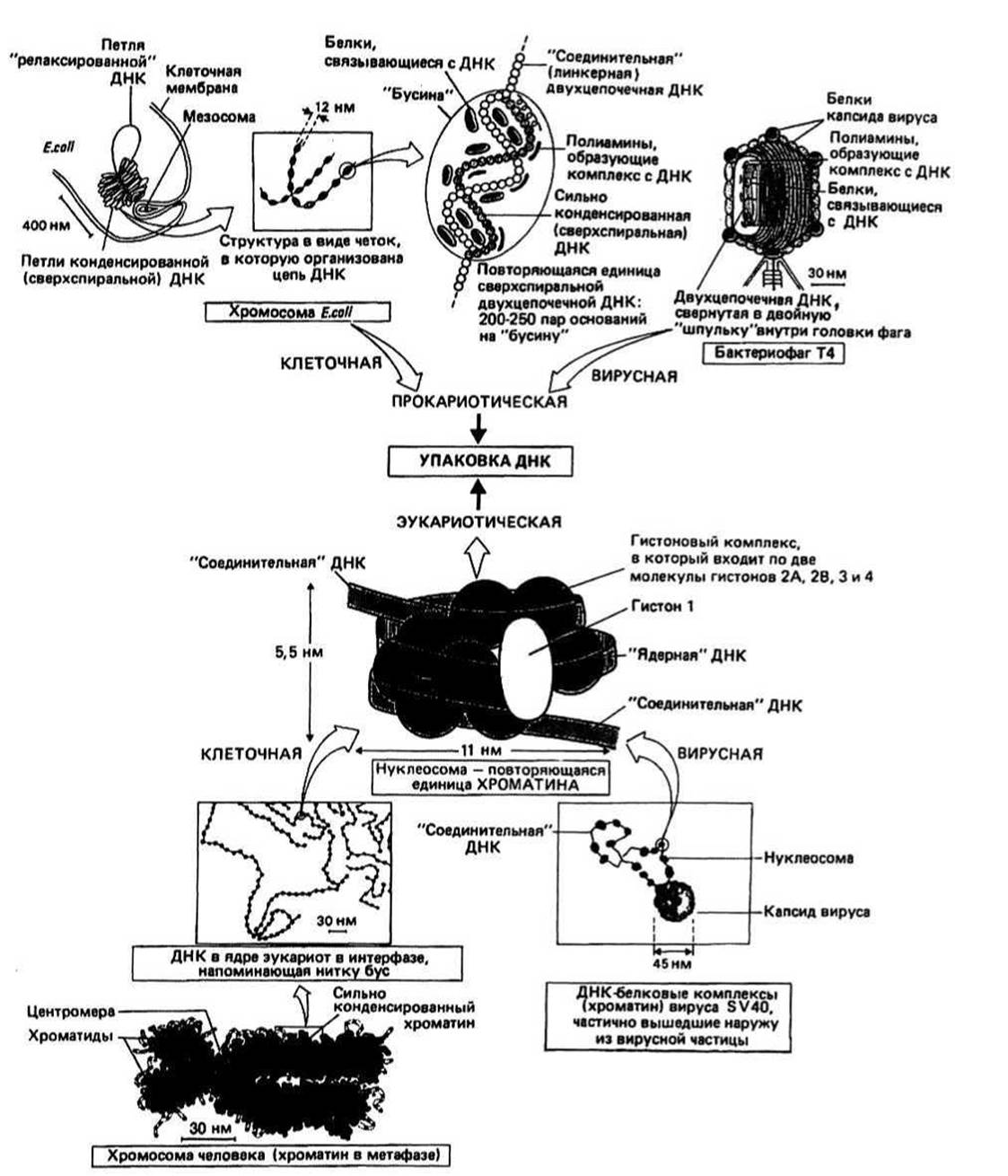
Рис.26.1.
Клетки прокариот содержат единственную копию геномной ДНК и являются, таким образом, гаплоидными. ДНК прокариотической клетки, кодирующая все клеточные белки и нуклеиновые кислоты (рРНК, тРНК и др.), входит в состав хромосомы. Е. coli, например, содержит единственную хромосому, являющуюся комплексом из ДНК, РНК и белков. ДНК представляет собой кольцевую молекулу и содержит 4,6 ∙ 106 пар нуклеотидов (или 4600 тысяч пар нуклеотидов, т.п.н.). Диаметр такого кольца составил бы примерно 1 мм, но, поскольку сама Е. coli имеет в поперечнике менее 2 мкм, ее ДНК должна быть плотно упакована (сконденсирована), чтобы поместиться внутри клетки.
Клетки эукариот устроены сложнее, чем клетки прокариот, и все они, за исключением гамет, содержат две совершенно одинаковые копии генома; другими словами, они диплоидны. Клетка нематоды содержит ДН К примерно в 40 раз, а клетка саламандры — в 40 000 раз больше, чем Е. coli. В клетке человека ДНК примерно в 700 раз больше, чем в Е. coli. Поскольку принято думать, что человек — это более сложный организм, чем саламандра, приходится заключить, что на основе суммарного содержания ДНК в клетке можно лишь весьма приблизительно судить о сложности того или иного организма.
ДНК эукариотических клеток находятся в ядре в виде набора отдельных фрагментов, называемых хромосомами. Каждая хромосома может содержать ДНК в количестве от 400 (дрожжи) до 100 000 т.п.н. (человек). Если бы всю клеточную ДНК в форме простой двойной спирали вытянуть в одну линию, то она имела бы слишком большую длину (1,74 м для клетки человека) и в таком виде не поместилась бы в ядре, поэтому хромосомы должны представлять собой сильно конденсированные структуры.
Вирусы могут содержать как одно-, так и двухцепочечную ДНК, которая может быть как непрерывной, так и состоящей из фрагментов. У некоторых вирусов (например, ВТМ, реовирусы; гл. 5) генетическим материалом служит РНК, которая также может быть как одно-, так и двухцепочечной.
Число генов в различных организмах можно оценить, исходя из того, что средняя длина гена составляет 1 т.п.н. В табл. 26.1 приведены соответствующие цифры.
Конденсация двухцепочечной ДНК, в результате которой продольные размеры молекулы уменьшаются в 10 000 раз, осуществляется одним из двух способов — путем сфероидальной намотки (т. е. свертывания в витки по сфероидной образующей) либо через образование сверхспиральной ДНК. Сфероидальная намотка по сути дела имеет место только в вирусах, тогда как образование сверхспиральной ДНК происходит во всех прокариотических и эукариотических клетках и во многих вирусах, поражающих эукариот.
Таблица 26.1
|
Организм |
Содержание ДНК (в парах нуклеотидов) |
Число генов |
|
SV40 (вирус животных) |
5,0 ∙ 103 |
5 |
|
Т4 (бактериофаг) |
2,0 - 105 |
200 |
|
Е. coli (бактерия) |
4,6 - 106 |
4600 |
|
Гаплоидная клетка человека |
2,8 - 109 |
100 000-500 000 |
Сфероидальная намотка ДНК происходит так, что в результате образуется компактная «шпулька». В бактериофаге Т4 ДНК свернута в двухслойную «шпульку» внутри головки фага, имеющей форму икосаэдра. Сначала образуются витки наружного слоя, затем — внутреннего; между слоями остается некоторый просвет. Оставшийся конец ДНК проходит через оба слоя и выводится в хвостовой отросток вируса через центральное отверстие (гл. 4). В других вирусах ДНК может свертываться не совсем так, как в Т4, но во всяком случае весьма похожим образом.
Сверхспиральная (сверхскрученная) ДНК образуется при введении в двойную спираль ДНК дополнительных витков (сверхвитков). В результате в молекуле возникает напряжение, которое проявляется, в частности, в том, что ось двойной спирали сама закручивается в спираль (спирализуется), что и показано на рис. 26.2 на примере кольцевой замкнутой двухцепочечной ДНК.
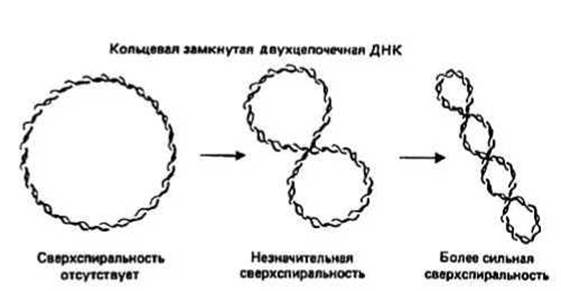
Рис. 26.2.
Стабилизация компактных форм ДНК необходима как в случае сфероидальной намотки ДНК, так и для сверхспиральной ДНК, поскольку в результате более плотной упаковки отрицательно заряженные фосфатные группы оказываются в непосредственной близости друг от друга. Взаимное отталкивание между этими группами компенсируется путем связывания их с положительно заряженными белками и с малыми молекулами, представляющими собой полиамины. Последние содержатся во всех без исключения прокариотических и эукариотических клетках, но отсутствуют во многих вирусах. К числу полиаминов, обнаруженных в эукариотических клетках, относится, например, спермидин, структура которого изображена на рис. 26.3. Специфические белки, стабилизирующие молекулу ДНК, есть в клетках любого типа, но лишь в немногих вирусах (например, в вирусе SV40).
1 По последним данным число генов у человека существенно меньше. — Прим. перев.
![]()
Рис. 26.3.
Упаковка ДПК в клетках прокариот имеет, вероятно, универсальный характер; покажем, как она осуществляется, например, в бактерии Е. coli. Сверхспиральный участок ДНК в комплексе с гистоноподобными (см. ниже) белками, обладающими сродством к ДНК, и полиаминами образует «бусину» диаметром около 12 нм. Внутри бусины ось двойной спирали ДНК перекручена и образует примерно шесть витков. Бусины соединены между собой участками ДНК, в которых сверхспиральность отсутствует, и объединяются, таким образом, в структуру, напоминающую ожерелье, четки или бусы. Такая организованная в бусы ДНК образует далее большие петли, которые стабилизируются и окончательно конденсируются благодаря взаимодействию с соответствующими белками и РНК.
Упаковка ДНК в эукариотических клетках включает в себя ассоциацию сверхспиральной ДНК с различными белками (гистонами), в результате которой образуется комплекс, называемый хроматином. Хроматин в свою очередь образует соленоидоподобную структуру, с которой связываются хромосомные структурные белки', получившийся в конечном счете комплекс называется хромосомой.
Хроматин состоит из двухцепочечной ДНК, которая обвивается вокруг особых гранул, состоящих из специфических белков — гистонов. Так образуется структура, похожая на бусы, которую называют хроматиновым волокном. Каждая бусина, называемая нуклеосомой, имеет около 10 нм в диаметре (например, в Е. coli). В результате конденсации ДНК в хроматин ее продольные размеры уменьшаются примерно в 5—6 раз.
Нуклеосома представляет собой сегмент двухцепочечной ДНК длиной около 200 пар оснований, навитый на белковую сердцевину, состоящую из восьми молекул белков — гистонов. Ее структура, впервые установленная Клугом с сотрудниками, изображена на рис. 26.1. Нуклеосомы обнаружены во всех эукариотических клетках и в некоторых вирусах, поражающих эукариот. Гистоны представляют собой основные белки с мол. массой (А/г) от 11 300 до 21 000. Всего известно пять типов гистонов: H1, Н2А, Н2В, НЗ и Н4. В нуклеосомную сердцевину входит по две молекулы гистонов Н2А, Н2В, НЗ и Н4. Обращенные наружу поверхности этих белковых молекул несут положительные заряды и образуют стабилизирующий остов, вокруг которого может закручиваться отрицательно заряженная молекула ДНК. Гистон HI размещается на участках ДНК, соединяющих одну нуклеосому с другой; ДНК этих участков называют поэтому соединительной (или линкерной) ДНК. Поскольку HI, по-видимому, не участвует в стабилизации структуры хромосомы, он, возможно, наряду с некоторыми другими белками, не входящими в число гистонов, играет какую-то роль в регуляции транскрипционной активности хроматина.
Соленоидоподобная структура клеточного хроматина была предложена для того, чтобы объяснить происхождение так называемых гетерохроматиновых волокон толщиной 20—30 нм, наблюдаемых в электронный микроскоп в интерфазном ядре (гл. 29). Каждый такой соленоид образуется в результате дальнейшей спирализации уже спирализованной дополнительно в области нуклеосом двойной спирали ДНК. На один виток соленоида приходится примерно 6 нуклеосом. Этот этап конденсации приводит к дальнейшему уменьшению продольных размеров ДНК — примерно в 40 раз по сравнению с размерами двухцепочечной ДНК. В настоящее время считают, что гетерохроматин представляет собой неактивную в отношении транскрипции форму хроматина. Поскольку продольные размеры ДНК в виде простой двойной спирали и той же ДНК в сильно конденсированной хромосоме различаются в 5 000—10 000 раз, осталось еще объяснить, откуда берется множитель 100-200 [40 ∙ 100(200) = 5 000(10 000)]. Пока, однако, неизвестно, как происходит дальнейшая конденсация соленоидоподобной структуры хроматина в его хромосомную форму.
Хромосомные структурные белки образуют каркас, на котором происходит окончательная конденсация хроматина в структуру с наиболее плотной упаковкой, характерной для эукариотических хромосом. Этот белковый остов не разрушается даже после удаления всех гистонов.