Введение в молекулярную биологию: От клеток к атомам - Энтони Рис, Майкл Стернберг 2002
Углеводы и липиды
Клеточная стенка бактерий
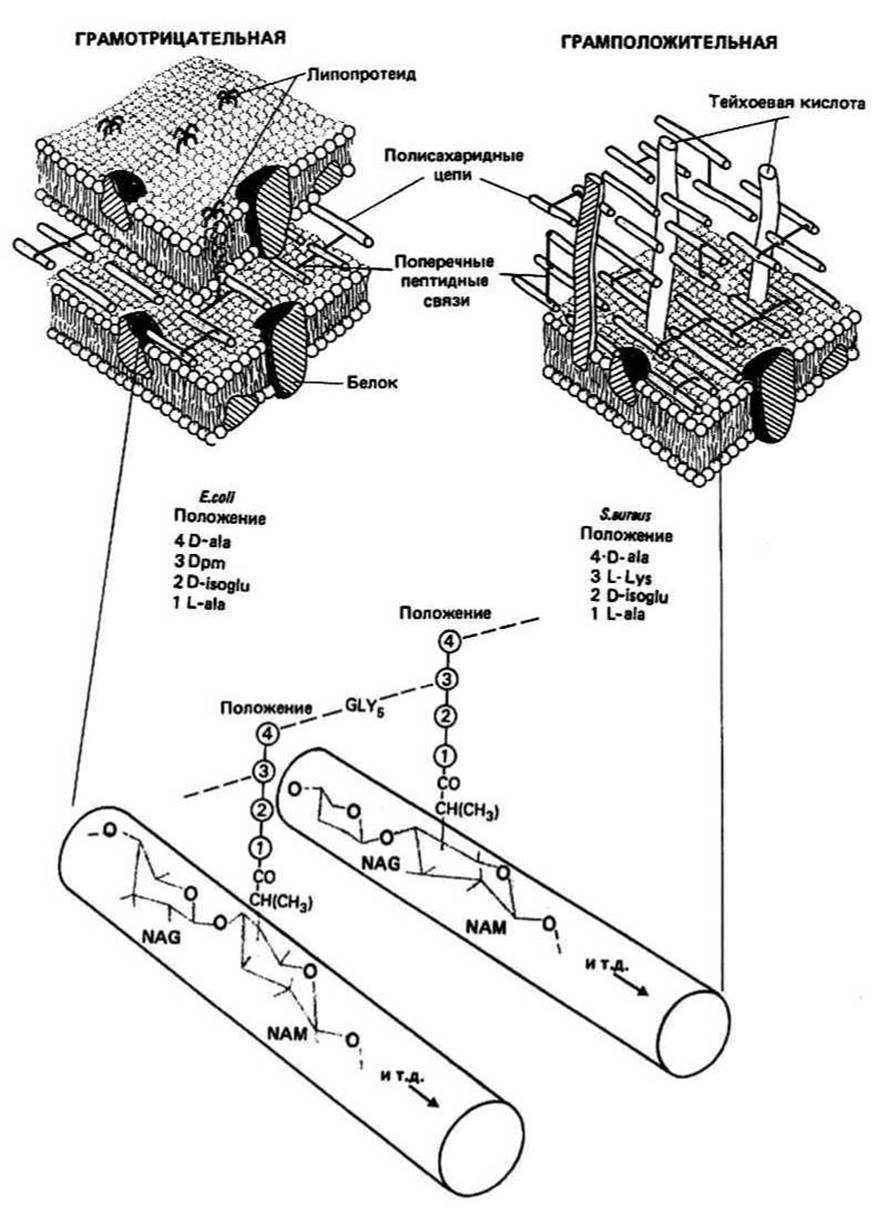
Рис. 35.1
Бактерии можно разделить на два класса в зависимости от их способности окрашиваться по методу, разработанному датским бактериологом Грамом в 1884 г. В процессе отмывки, которая следует за процедурой окрашивания, грамположительные бактерии удерживают фиолетовый краситель, а грамотрицательные теряют его. Только в 60-х годах Сэлтон и другие показали, что это явление обусловлено фундаментальными различиями в структуре клеточных стенок двух указанных классов бактерий. Морфологические исследования, выполненные с помощью электронного микроскопа, показали, что грамположительные бактерии обладают однослойной электроноплотной нелипидной клеточной стенкой толщиной 20—80 нм, окружающей плазматическую мембрану толщиной 7,5 нм. У грамотрицательных же бактерий клеточная стенка двуслойная. Первый слой — очень тонкая (толщиной 1 нм) нелипидная мембрана — окружен вторым: липидной мембраной толщиной 7,5 нм. Исследования мембран грамположительных и грамотрицательных бактерий методом замораживания—скалывыния подтвердили, что у них имеется два и три слоя соответственно.
Клеточная стенка — это жесткая структура, которая служит для поддержания формы клетки и предотвращает ее лизис из-за осмотического шока. При обработке бактерий гидролитическим ферментом, таким, как лизоцим (гл. 14), клеточная стенка может «раствориться». В случае грамположительных бактерий после такой обработки остаются клетки, чувствительные к осмотическому шоку, имеющие только плазматическую мембрану и называемые протопластами. У грамотрицательных бактерий вторая, наружная, мембрана сохраняется и образующиеся клетки, называемые сферопластами, несколько менее чувствительны к осмотическому шоку. Материал клеточных стенок, который теряется в обоих случаях, известен как пептидогликан. Его также называют гликопептидом или мукопептидом.
Пептидогликан — это сложный матрикс, содержащий полисахаридные цепи, связанные друг с другом поперечными сшивками из коротких пептидных цепей. Когда гидролиз клеточной стенки бактерий любого типа проводят до расщепления ее на мономерные составляющие, единственными образующимися моносахаридами являются N-ацетилглюкозамин (NAG) и N-ацетил мурамовая кислота (NAM). ОН-группы обоих моносахаридов, связанные с С-атомом, имеют (3-конформацию. Полисахарид пептидогликан — это Р(1 → 4)-цепь из чередующихся Сахаров NAG-NAM. У грамположительных бактерий (например, у Bacillus subtilis) эти цепи образуют трехмерный волокнистый матрикс, имеющий до 40 слоев, в то время как у грамотрицательных (например, у Е. coli) слой только один. Таким образом, толщина пептидогликанового матрикса может составлять от 1 до 80 нм. Полисахаридные цепи связаны друг с другом пептидными цепочками. Состав пептидных поперечных сшивок был установлен с помощью кислотного гидролиза очищенного материала клеточных стенок. Были обнаружены как L-, так и D-аминокислоты. Реконструкция поперечных связей показала, что они содержат два типа структур. Во-первых, это тетрапептидный элемент, который присоединен к каждой молекуле сахара NAM, во-вторых, пептидный мостик, который соединяет между собой полисахаридные цепи через посредство тетрапептидных элементов.
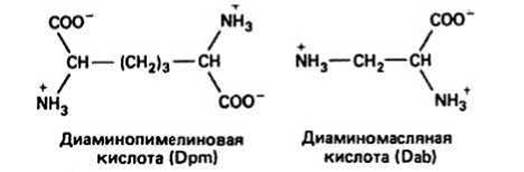
Рис. 35.2.
Тетрапептидные элементы различаются по своему составу. Они присоединены к сахару NAM через СООН-группу атома 3С. Первой аминокислотой в пептиде всегда является L-аланин (положение 1). Следующим остатком может быть D-изоглутаминовая кислота (приставка «изо» означает, что в образовании пептидной связи участвует СООН-группа боковой цепи, а группа сс-СООН основной цепи остается свободной) или какой-нибудь минорный остаток (положение 2). Третий остаток, как правило, содержит в боковой цепи аминогруппу. Обычно им является L-лизин, хотя это могут быть и L-диаминопимелиновая (Dpm) или L-диаминомасляная (Dab) кислоты (рис. 35.2). Последним остатком всегда является D-аланин (положение 4). Таким образом, тетрапептид представляет собой чередующуюся цепь из L- и D- аминокислот. Это способствует большей устойчивости его к протеолитическому расщеплению и соответственно большей жизнестойкости бактерий. Полная структура этого элемента дана на рис. 35.3. Таким образом, каждый тетрапептидный элемент одной цепи NAG—NAM связан в поперечном направлении с другими цепями NAG—NAM либо по третьему, либо по четвертому положению. Чтобы сохранилась симметрия клеточной стенки, эти связи образуются между положением 3 тетрапептидного элемента одной цепи и положением 4 другой. Так образуется непрерывная сеть поперечных связей.
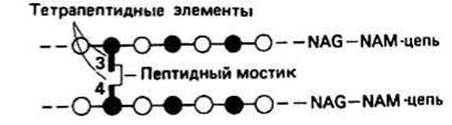
Рис. 35.3.
Пептидный мостик обычно представляет собой пентапептидную цепочку, состоящую только из аминокислот с короткими боковыми цепями. Типичным мостиком является пентаглицин, хотя встречаются и мостики L-Ala4— L-Thr и Gly3— L-Ser2. Ковалентные связи образуются на одном конце между СООН-груп- пой мостика и МН2-группой боковой цепи остатка в положении 3, а на другом — между NH2-rpyппой мостика и а-СООН-группой D-аланина в положении 4. Иногда бактериальная стенка оказывается весьма чувствительной к гидролитическим ферментам, таким, как лизоцим (гл. 14). Это может быть обусловлено ее более открытой структурой из-за наличия особых пептидных мостиков, которые формируются из тетрапептидных элементов, соединенных конец к концу, так что при этом образуются длинные поперечные связи. Для появления межцепочечных связей такого типа должна быть удалена часть тетрапептидных элементов с некоторых Сахаров NAM. В результате в целом число поперечных связей уменьшается и полисахаридные цепи оказываются более доступными для гидролаз. Подходящим субстратом для исследования активности лизоцима является препарат клеточной стенки бактерий типа Micrococcus lysodeikticus. Сложные углеводные цепи имеются только у грамположительных бактерий. Это тейхоевые и липотейхоевые кислоты.
Тейхоевые кислоты — это цепочки из молекул глицерола или рибитола, связанных друг с другом фосфодиэфирными мостиками. На одну цепь может приходиться до 30 молекул. Кроме того, в цепочку могут быть включены остатки Сахаров или аминокислот. На рис. 35.4 показана тейхоевая кислота из S. aureus, построенная на основе рибитола. У некоторых видов бактерий молекулы тейхоевой кислоты, расположенные перпендикулярно поверхности плазматической мембраны, соединены с пептидогликаном через ОН-группу при атоме 6С остатков NAM. Таким образом, слои пептидогликана оказываются связанными друг с другом.
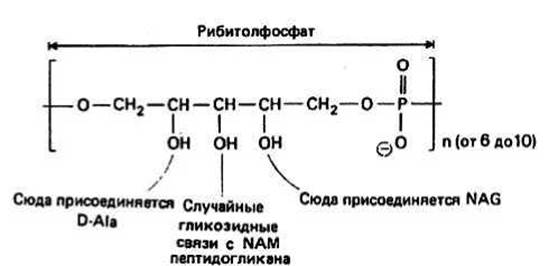
Рис. 35.4.
Липотейхоевые кислоты подобны тейхоевым, но связаны не с пептидогликаном, а с гликолипидными молекулами, погруженными в плазматическую мембрану.
Механизм действия некоторых антибиотиков состоит в их «вмешательстве» в сборку клеточной стенки. Так, пенициллин блокирует фермент, в функцию которого входит образование поперечных связей между тетрапептидами и пептидными мостиками в реакции транспептидации (гл. 42).