Биологические мембраны - А. Н. Огурцов 2012
Структура и функции биомембран
Мембранные липиды
Латеральная мобильность в мембранах
Тепловое движение липидных молекул сочетает в себе практически свободное вращение относительно оси молекулы и латеральную диффузию в плоскости мембраны. Гидрофобные жирнокислотные хвосты липидов при таких двух типах движения остаются погружёнными в гидрофобный внутренний слой мембраны.
И в природных, и в искусственных модельных мембранах частота латеральных перескоков липидов при физиологической температуре 37°С порядка 107 с-1 и молекула пробегает за секунду расстояние порядка нескольких микрометров. Такая скорость латеральной диффузии свидетельствует о том, что вязкость биомембраны в 100 раз больше вязкости воды и приблизительно равна вязкости растительного масла. И, несмотря на то, что молекулы липидов диффундируют в биомембране медленнее, чем компоненты окружающего водного раствора, любая молекула мембранного липида пробегает расстояние, сравнимое с длиной бактериальной клетки (~1 мкм) за 1 секунду, а расстояние порядка длины животной клетки за 20 секунд.
Абсолютные значения коэффициентов латеральной диффузии липидов и белков в биомембране можно определить методом восстановления флуоресценции после фотообесцвечивания (fluorescence recovery after photobleaching, FRAP). Исследуемые мембранные объекты, например белки определённого сорта, предварительно маркируют флуоресцентными метками (рисунок 20 (1)).
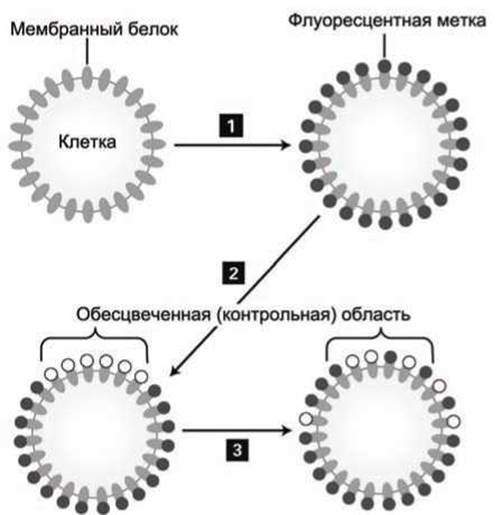
Рисунок 20 - Последовательность операций в методике FRAP
Затем лазерным лучом необратимо разрушают флуоресцентные метки в пределах определённой контрольной области, что приводит к резкому снижению интенсивности флуоресценции из этой области (рисунок 20(2)). С течением времени подвижные белки диффундируют за пределы этой области, а на их место приходят белки с необеспеченными метками, что приводит к росту интенсивности флуоресценции от контрольной области мембраны (рисунок 20(3)).
Если часть данных белков неподвижно закреплена в мембране, то уровень флуоресценции после выхода кривой на насыщение будет ниже, чем до обесцвечивания, что позволяет определить относительные доли подвижных и неподвижных маркированных молекул. На рисунке 21 приведён пример такого рода кривой для случая, когда 50% меченых белков были неподвижными.
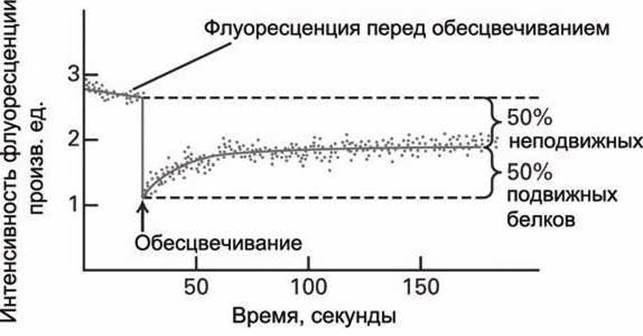
Рисунок 21 - Восстановление флуоресценции после фотообесцвечивания
Из такой кинетической кривой может быть получено численное значение коэффициента диффузии маркированного белка в мембране.
В результате исследований методом FRAP было показано, что в плазматических мембранах фибробластов (основные клетки соединительной ткани позвоночных, синтезирующие коллаген) все фосфолипиды могут свободно перемещаться только на расстояние порядка 0,5 мкм, но не больше. По-видимому, мембрана фибробласта мозаично разбита на белковые и липидные "острова" размером порядка 1 мкм, и липиды могут свободно перемещаться только в пределах своего острова. Кроме того, оказалось, что коэффициент диффузии липидов в плазматической мембране в десять раз ниже, чем в "чистом" липидном бислое (1(TS см2/с и 10-7 см2/с, соответственно). На основании этих фактов можно сделать вывод, что липиды могут быть ассоциированы (обратимо связаны) с определёнными мембранными белками.