Биологические мембраны - А. Н. Огурцов 2012
Структура и функции биомембран
Пассивный трансмембранный транспорт
Диффузия молекул воды через мембрану
В фосфолипидной фазе мембраны хорошо растворимы неполярные вещества, например органические жирные кислоты, эфиры. Эти вещества хорошо проникают через липидную фазу мембраны. Плохо проходят через липидный бислой полярные, водорастворимые вещества: соли, основания, сахара, аминокислоты, спирты.
На этом фоне представляется необъяснимым аномально большое значение коэффициента проницаемости липидной мембраны для воды. В последнее время проникновение через липидные бислойные мембраны мелких полярных молекул связывают с образованием между жирнокислотными хвостами фосфолипидных молекул при их тепловом движении небольших свободных полостей — кинков (от англ. kink - петля), образованных гош-транс-гош-конфигурацией липидных молекул (рисунок 50).
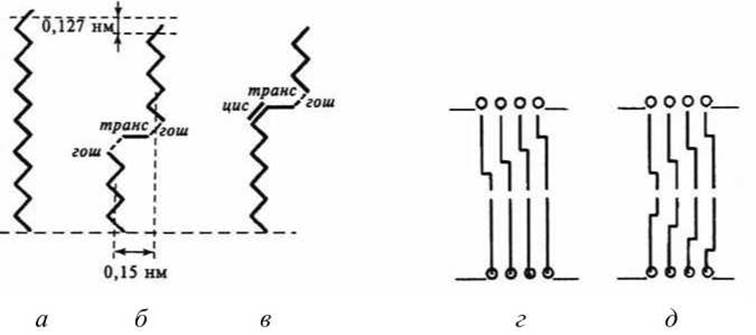
Рисунок 50 - Схема углеводородных цепей в биомембране: а - в полностью транс-конфигурации; б - в гош-транс-гош-конфигурации; в - в цис-транс-гош-конфигурации. Кинк-блоки в углеводородных цепях мембран: г - в одном полуслое; д — в двух слоях липидного бислоя
Гош-конформации (гош (+) и гош (-), поворот на ±120° относительно транс-конформации) сравнительно мало превышают по энергии транс-конформацию (на 2-3 кДж/моль), но эти состояния разделяет энергетический барьер высотой 12-17 кДж/моль. Если углеводородные цепи в полностью транс-конфигурации представляют собой линейные структуры, то появление одиночной гош-конформации в цепи приводит к искривлению пространственной конфигурации цепи на угол -120°.
В плотно упакованных мембранных системах с полностью транс-конформацией углеводородных цепей это искривление порождает серьезные стерические затруднения, делающие невозможным появление одиночных гош-конформаций.
Уменьшение стерических затруднений при плавлении углеводородных цепей в мембранах достигается при синхронном появлении в цепи сразу двух гош-конфигураций (гош (+) и гош (-)), разделённых С-С-связью в транс-конформации.
Хотя энергия такого состояния вдвое выше одиночной гош-конформации, возникающее ротационное состояние цепи не вызывает сильного расширения решётки, так как при последовательном повороте цепи на +120 и -120° пространственная конфигурация цепи в целом сохраняется прямолинейной (рисунок 50). Участок цепи, находящийся в гош(+)-транс-гош(-)-конформации, формирует уступ или петлю в углеводородной цепи - кинк.
Образование кинка сопровождается уменьшением эффективной длины цепи на ~ 0,127 нм. При этом часть цепи отодвигается на ~ 0,15 нм, образуя свободный объём, а занимаемый молекулой липида общий объём увеличивается на 0,025-0,050 нм3 (рисунок 50).
Хотя появления одного кинка в углеводородной цепи недостаточно для её плавления, однако одиночные кинки облегчают возникновение кинков в соседних углеводородных цепях, формируя чередующиеся кинк-блоки (рисунок 50(г)). Такие блоки могут возникать либо в одном полуслое мембраны, либо в двух противоположно расположенных углеводородных цепях. При увеличении числа кинков в углеводородных цепях разупорядоченностъ углеводородной зоны мембран резко нарастает.
Вследствие теплового движения хвостов кинки могут перемещаться через мембрану и переносить попавшие в них мелкие молекулы, в первую очередь, молекулы воды. Двойные (цис-) связи в жирнокислотных ненасыщенных цепях (рисунок 50(b)) мембран могут играть роль инициаторов образования кинков в соседних насыщенных цепях. В этом случае для образования кинка в ненасыщенной цепи необходимо появление лишь одной гош-конформации при искривлении цепи на 80°.
При этом устраняются стерические затруднения, возникающие при размещении ненасыщенной цепи в углеводородной зоне мембран из насыщенных липидов, что хорошо согласуется с экспериментально наблюдаемым резким снижением температуры ТФП фазового перехода мембран из насыщенных липидов при добавлении к ним небольших количеств ненасыщенных жирнокислотных цепей.
Таким образом, низкую вязкость углеводородной зоны мембран в жидкокристаллическом состоянии (при Т > ТФП) связывают с
1) возрастанием амплитуды крутильных осцилляций вокруг С-С связей;
2) появлением гош-конформаций (кинки);
3) их быстрой изомеризацией в соседние положения.
Для оценки эффективности этих процессов сравним частоту крутильных колебаний вокруг С-С связей 8-1012с-1 с частотой возникновения гош-конформаций при комнатной температуре.
Полагая, что величина барьера, разделяющего транс-гош-конформации, равна 12 кДж/моль, находим
![]()
Отсюда следует, что гош-конформации при комнатных температурах возникают с высокой частотой вследствие крутильных осцилляций.
В условиях высокой текучести гидрофобной зоны мембран кинк может изомеризоваться, смещаясь вдоль углеводородной цепи за счёт синхронного поворота на 120° соответствующей последовательности С-С связей. При смещении в соседнее положение кинк передвигается на ~13 нм. Такое перемещение кинка вдоль углеводородной цепи можно рассматривать как своеобразную одномерную диффузию, которую можно охарактеризовать коэффициентом диффузии DK =0,5vK(∆L)2, где vK - частота скачка кинка; ∆L - шаг одного скачка. Полагая, что частота скачка кинка по порядку величины соответствует частоте появления гош-конформаций, находим DK≈ 10-5см2/с.
Полученное значение практически совпадает с известными коэффициентами проницаемости липидных мембран для кислорода, воды и небольших молекул неэлектролитов. Это совпадение, а также геометрическое соответствие размеров таких молекул и свободного объёма, возникающего при образовании кинка, позволяет утверждать, что трансмембранный перенос малых молекул осуществляется внутри свободного объёма, образуемого кинком.