Биологические мембраны - А. Н. Огурцов 2012
Структура и функции биомембран
Активный транспорт
Протонные насосы V-класса
Все АТФазы V-класса переносят через мембрану только протоны (ионы Н+). Функционирование эти протонных помп, расположенных на мембранах лизосом, эндосом и вакуолей растений, обеспечивает повышенную кислотность в люмене этих органелл, которая может в сотни раз (значения pH ~ 4,5-5,0) превышать кислотность цитоплазмы (pH ~ 7,0).
АТФазы V-класса структурно подобны насосам F-класса (рисунок 63(a)), они состоят из двух доменов: цитозольного гидрофильного домена V1 и трансмембранного домена V0, каждый из которых состоит из множества субъединиц (рисунок 71).
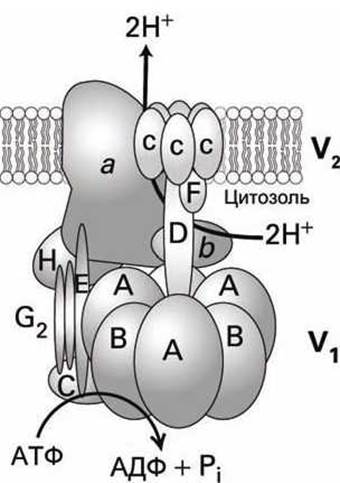
Рисунок 71 - Схема АТФ-насоса V-класса
Связывание и гидролиз АТФ на субъединице В домена V1 обеспечивает энергией процесс перекачки протонов через канал, образуемый субъединицами а и С в мембранном домене V0 (рисунок 71).
В отличие от Р-насосов, насосы класса V не фосфорилируются в процессе функционирования. А в отличие от F-насосов, насосы класса V не используются для синтеза АТФ, а всегда переносят протоны из цитозоля в люмен органеллы. Для существенного изменения кислотности оказывается достаточным транспортировать относительно немного протонов через мембрану. Действительно, для сферической лизосомы диаметром 0,2 мкм (соответственно, объёмом 4,18-10-15 см3) кислотность pH = 4 обеспечивается всего лишь 252 протонами на всю лизосому.
Самостоятельно АТФазы V-класса не способны значительно повысить кислотность люмена органеллы, поскольку они являются электрогенными насосами, которые осуществляют процесс разделения зарядов на мембране. Действительно, закачанные ионы Н+ в люмен органеллы продолжают электростатически взаимодействовать с "оставшимися" в цитозоле анионами (например, ОН= или Сl-), что приводит к тому, что разделённые мембраной заряды концентрируются на этой же мембране (рисунок 72(a)). В результате на мембране создаётся электрический (мембранный) потенциал, а в люмене органеллы не оказывается избыточных протонов, и кислотность люмена остаётся нейтральной.
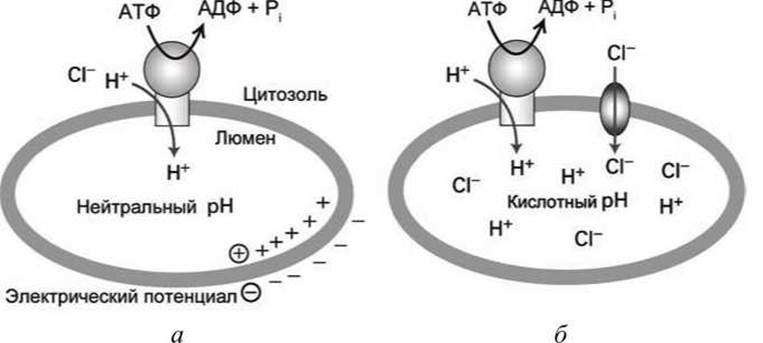
Рисунок 72 - Функционирование АТФаз V-класса: а - создание мембранного потенциала, б - повышение кислотности люмена органеллы
По мере увеличения мембранного потенциала возрастает электростатическое противодействие перемещению протонов через мембрану, которое в конце концов останавливает перенос протонов V-насосами задолго до того, как кислотность люмена органеллы заметно увеличится.
Именно таким способом протонные помпы P-класса на плазматических мембранах клеток растений и дрожжей создают отрицательный потенциал цитозоля этих клеток.
Для того чтобы повысить кислотность люмена органеллы или внеклеточного пространства (например, люмена кишечника), перемещение протонов через мембрану должно сопровождаться либо (1) синхронным перемещением в том же направлении эквивалентного количества анионов (например, Сl-), либо (2) синхронным перемещением каких-либо других катионов навстречу току протонов, сохраняя тем самым неизменным объёмный электрический заряд компартмента.
Процесс первого типа осуществляется в лизосомах и вакуолях растений, чьи мембраны наряду с АТФазами V-класса содержат также анионные каналы, по которым синхронно (и самопроизвольно) перемещаются ионы Сl- внутрь люмена органеллы (рисунок 72(6)).
Процесс второго типа осуществляется на апикальных поверхностях плазматических мембран клеток слизистой оболочки кишечника (рисунок 22), которые содержат Р-насосы Н+/К+ АТФазы, которые работают по схеме антипорта и не являются электрогенными, поскольку за один цикл переносят через мембрану клетки один протон из цитозоля наружу в люмен кишечника и один ион калия К+ из люмена кишечника внутрь клетки.