Биологические мембраны - А. Н. Огурцов 2012
Электрогенез биомембран
Электрические потенциалы биомембран
Потенциал покоя
Мембранные потенциалы подразделяются на потенциалы покоя и потенциалы действия.
Потенциалом покоя (resting voltage) называют стационарную разность электрических потенциалов, которую можно зарегистрировать между внутренней и наружной поверхностями мембраны в невозбуждённом состоянии. Поскольку биомембраны разделяют изолированные друг от друга компартменты, то потенциал покоя определяется двумя факторами:
1) различием в концентрации ионов по разные стороны мембраны;
2) диффузией ионов через мембрану.
Величина мембранного потенциала покоя составляет порядка 70 мВ, причём цитозоль клетки всегда заряжен отрицательно по отношению к положительно заряженной внеклеточной среде. Учитывая малую толщину биомембраны (порядка 3,5 нм) градиент потенциала на мембране (напряжённость электростатического поля) составляет 200 000 В/см.
И градиент концентрации ионов через мембрану, и мембранный потенциал играют определяющую роль во множестве биологических процессов. Так, рост цитозольной концентрации Са2+ является важным управляющим сигналом, который, например, инициирует сокращение клеток мускулатуры или запускает процессы секреции пищеварительных ферментов в клетках поджелудочной железы. Во многих животных клетках совместный эффект градиента концентрации ионов Na+ и мембранного потенциала индуцирует трансмембранное перемещение аминокислот и других молекул против их градиента концентрации по механизму симпорта или антипорта. Ещё один пример - распространение нервного импульса по нейронам определяется работой ионных каналов.
Для наглядности рассмотрим модельную систему, в которой два электролита с концентрациями ионов, соответствующими физиологическим концентрациям ионов калия и натрия в цитозоле и экзоплазме, разделены мембраной, которая не проницаема для ионов калия и натрия (рисунок 89). Электрический потенциал на такой мембране равен нулю.
Если концентрация какого-либо иона внутри клетки свн отлична от концентрации этого иона снаружи снар, и мембрана проницаема для этого иона, возникает поток заряженных частиц через мембрану, вследствие чего нарушается электрическая нейтральность системы, образуется разность потенциалов внутри и снаружи клетки φМ = φвн - φнар, которая будет препятствовать дальнейшему перемещению ионов через мембрану.
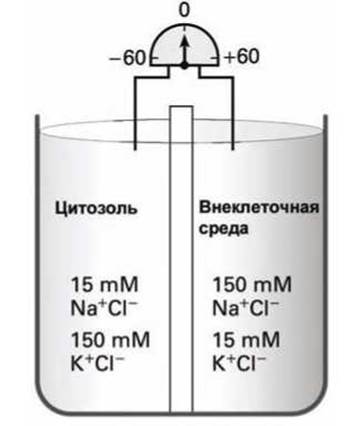
Рисунок 89 - Схема, моделирующая ионный состав цитозоля и экзоплазмы
При установлении равновесия выравниваются значения электрохимических потенциалов по разные стороны мембраны ![]()
Так как
![]()
![]()
Другой вид этого равенства, в котором в левую часть вынесен только мембранный потенциал φМ = φвн - φнар, называется формула Нернста для равновесного мембранного потенциала
![]()
Если мембранный потенциал обусловлен переносом ионов К+, для которого [K+]вн > [К+ ]нар и z = +1, то равновесный мембранный потенциал отрицателен и имеет вид
![]()
Для ионов Na+: [Na+]вн < [Na+]нap, z = +1,
![]()
Для ионов Cl-: [Cl-]вн < [Cl-]нap, z = -1,
![]()
При T =300 К и z = +1 (с учётом ln х = 2,3 lg х)
![]()
откуда следует
![]()
Величина мембранного потенциала покоя большинства клеток практически совпадает с равновесным мембранным потенциалом для ионов калия. Исходя из этого ещё в 1902 году Бернштейн предположил, что причиной мембранного потенциала покоя является диффузия ионов калия из клетки наружу через какие-то каналы в мембране.
Это предположение было впоследствии подтверждено экспериментально, и были обнаружены и исследованы мембранные белки, которые формируют каналы в мембранах, селективно пропускающие через себя различные ионы.
Действительно, в случаях, когда в мембране присутствуют селективные ионные каналы для ионов натрия (рисунок 90(a)) или калия (рисунок 90(6)), то соответствующее перемещение ионов под действием градиента концентрации приводит к разделению зарядов на мембране, в результате чего на мембране формируется мембранный потенциал.
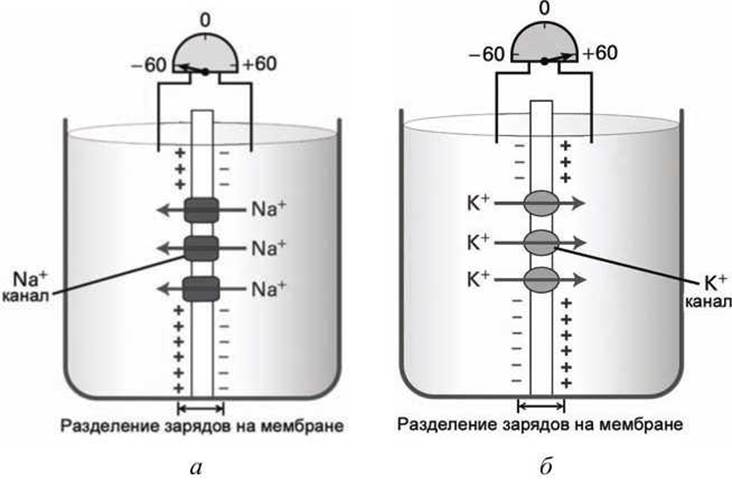
Рисунок 90 - Роль селективных ионных каналов в формировании мембранного потенциала: а - натриевые каналы, б - калиевые каналы
Для случая концентрации ионов натрия и калия, указанных на рисунке 83 мембранный потенциал за счёт перемещения ионов натрия через мембрану будет равен φ = -59 мВ (рисунок 90(a)), а в случае селективного трансмембранного транспорта ионов калия через мембрану, потенциал мембраны будет равен φ = +59 мВ (рисунок 90(6)).
Экспериментально измеренные значения мембранных потенциалов и концентраций ионов, а также рассчитанные по формуле Нернста значения потенциалов для клеток двух типов представлены в таблице 5.
Таблица 5 - Содержание ионов К+, Na+, Cl-, равновесные потенциалы φ0М и потенциалы покоя φеМ некоторых клеток
|
Объект |
Концентрация, ммоль/л |
φM, мВ, по формуле Нернста |
φeM, мВ, экспер. |
|||||||
|
[К+] |
[Na+] |
[Сl-] |
К+ |
Na+ |
Cl- |
|||||
|
вн |
нар |
вн |
нар |
вн |
нар |
|||||
|
Гигантский аксон |
360 |
10 |
70 |
420 |
160 |
500 |
-90 |
+50 |
-зо |
-60 |
|
Мышца лягушки |
125 |
2,5 |
15 |
125 |
и |
120 |
-98 |
+60 |
-87 |
-94 |
Пользуясь формулами электростатики, оценим, какое количество ионов должно перейти из цитоплазмы во внеклеточную среду, чтобы создать разность потенциалов порядка 10-1 В.
Радиус клетки равенrг = 10 мкм= 10-5м. Удельная электроёмкость мембраны (электроёмкость на единицу площади) равна Cs = 10-2 Ф/м2.
Площадь мембраны составляет S = 4πr ≈ 4π10-10 м2≈ 10-9 м2. Тогда электроёмкость мембраны будет С = Cs∙ S ≈ 10-2 Ф/м2 ∙ 10-9м2 = 10-11Ф.
Величина заряда каждого знака на поверхности мембраны, если представить её в виде конденсатора, |q| = Сφ ≈ 10-11∙ 10-1 = 10-12Кл, что соответствует
![]()
моль ионов.
Объём клетки данного радиуса
![]()
Изменение концентрации ионов в клетке вследствие выхода из клетки 10-17 моль ионов составит
![]()
Это изменение концентрации ничтожно по сравнению с концентрацией ионов калия внутри клетки (таблица 5), оно составляет всего 10-4 % от внутриклеточной концентрации калия.
Таким образом, чтобы создать равновесный нернстовский мембранный потенциал, через мембрану должно пройти пренебрежимо малое количество ионов по сравнению с общим их количеством в клетке.