Биологические мембраны - А. Н. Огурцов 2012
Электрогенез биомембран
Вторичный транспорт котранспортёрами
Транспортёры в вакуолях растений
Кислотность люмена вакуоли (pH = 3 : 6) выше, чем у цитозоля растений (pH = 7,5). Кислотность вакуоли поддерживается с помощью АТФ-насосов V-типа и специфических для растений РРi-насосов, использующих энергию гидролиза пирофосфата. Оба эти насоса располагаются на мембране вакуоли и импортируют протоны внутрь вакуоли против градиента концентрации.
Мембрана вакуоли содержит также каналы для Сl- и NO-3, которые пропускают эти анионы из цитозоля в люмен вакуоли. Стимулирующим фактором для перемещения этих анионов является положительный электрический потенциал, который создаётся протонами, закачиваемыми в вакуоль протонными помпами (рисунок 105).
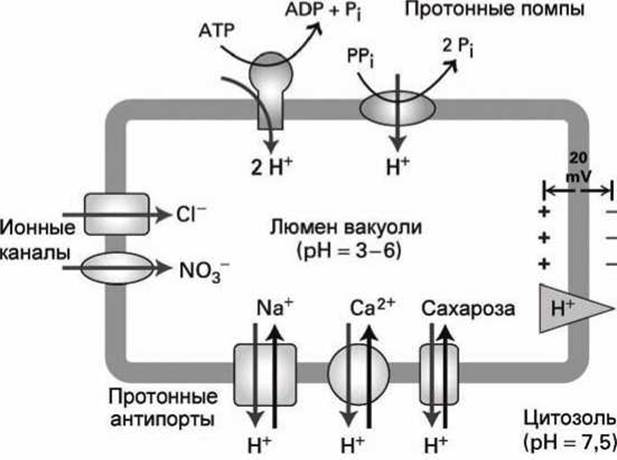
Рисунок 105 - Транспортные системы вакуоли клетки растений
Комбинированное действие протонных помп и анионных каналов поддерживает мембранный потенциал мембраны вакуоли 20 мВ и повышенную кислотность люмена. Протонный градиент на мембране вакуоли используется (аналогично градиенту ионов натрия на плазматической мембране) в качестве источника энергии для работы котранспортёров.
На рисунке 105 показаны три таких протонных антипортёра, которые используют энергию тока протонов из вакуоли в цитозоль (по градиенту концентрации и под действием избыточного положительного заряда люмена вакуоли) для закачивания в вакуоль катионов натрия и кальция, а также молекул сахарозы. Таким способом избыток сахарозы, который создаётся в листьях днем в результате фотосинтеза, запасается в вакуолях клеток. Ночью сахароза переносится в цитоплазму и метаболизируется до СО і и Н2O, в ходе чего синтезируется АТФ из АДФ и Рi.