Биологические мембраны - А. Н. Огурцов 2012
Электрогенез биомембран
Внутриклеточная сигнализация
Сигнальная функция биомембран
В плазматической мембране клетки находятся молекулярные комплексы, преобразующие внешние сигналы во внутриклеточные. Рецепторы распознают сигнальные молекулы и приводят в действие внутриклеточные пути передачи информации, которые, в конечном счёте, ведут к регуляции клеточных процессов, например, секреции, сокращению, метаболизму или росту.
Внутриклеточные сигналы распространяются внутри клетки молекулами-посредниками - вторичными месенджерами (second messengers, от англ. messenger - посыльный).
На молекулярном уровне процесс передачи информации через мембрану обеспечивается цепочкой мембранных белков, последовательно взаимодействующих друг с другом.
Каждое очередное взаимодействие вызывает конформационную перестройку следующего в цепочке белка - изменение его структуры, а, следовательно, и функции.
Молекулярные машины, обеспечивающие передачу сигнала от рецепторов к внутриклеточным мишеням, состоят, как правило, из нескольких белковых компонентов, совокупность которых обычно называют каскадом передачи сигнала или просто каскадом.
Белковые компоненты каскадов передачи сигнала называют белковыми посредниками или медиаторами.
На определенной стадии каскада дальнейшая передача информации поручается находящимся в цитоплазме малым молекулам или даже ионам. Они-то и являются вторичными посредниками. Их диффузия обеспечивает быстрое распространение сигнала по всей клетке.
Для обозначения сигнальных молекул принято использовать термин месенджер, а не посредник. Дело в том, что в цитоплазме в передачу сигнала вовлечены как разнообразные белки, так и малые молекулы (собственно вторичные сигналы), причём функционально все они являются посредниками (медиаторами) между рецептором, на который подействовал внешний стимул, и клеточным ответом.
Однако между ними есть и принципиальное различие: белки образуют своеобразную молекулярную машину, которая, с одной стороны, чувствует внешний сигнал, а с другой - обладает ферментативной или иной активностью, модулируемой этим сигналом, в то время как малые молекулы действительно служат месенджерами (курьерами, посыльными, гонцами) между различными белками, полиферментными комплексами или даже клеточными структурами.
Наиболее часто используемые месенджеры:
✵ циклический аденозин монофосфат (цАМФ);
✵ циклический гуанозин монофосфат (цГМФ);
✵ инозитол трисфосфат (ІР3);
✵ диацилглицерол (ДАГ);
✵ катион кальция (Са2+).
Число различных вторичных месенджеров оказалось удивительно небольшим. Иными словами, пути передачи внутриклеточных сигналов универсальны и способны регулировать множество разнообразных физиологических и биохимических процессов.
Выделяют два основных пути передачи сигналов.
В одном из них вторичным месенджером служит циклический аденозин монофосфат (цАМФ).
В другом - действует комбинация трёх вторичных посредников: ионы кальция, инозитол трисфосфат и диацилглицерол. Последние два вещества образуются в самой плазматической мембране.
Эти два пути передачи сигналов имеют много общего. На рисунке 138 показана обобщённая схема мембранной рецепции химических сигналов, основные этапы которой присутствуют в мембранных сигнальных механизмах, существующих в клетках разного типа.
В обоих случаях элементы, получающие информацию от первого звена - рецептора, и передающие её через мембрану внутрь клетки, представляют собой так называемые G-белки - мембранные белки-ГТФазы, активирующиеся при связывании гуанозинтрифосфата.
G-белки активируют усилительный фермент, находящийся внутри клетки. А фермент уже превращает молекулы вещества-предшественника в молекулы вторичного месенджера.
Работу мембранного конвейера можно представить в виде последовательных четырёх этапов.
Первый этап (рисунок 138(а,б)). Сигнальная молекула, например, гормон (1) (рисунок 138(a)), по кровотоку или лимфатической системе доставляется к рецептору GPCR (G-protein coupled receptor) (2).
Присоединение гормона к рецептору вызывает трансформацию последнего, изменяющую конформацию внутриклеточной части рецепторного белка, что превращает его в фактор замены нуклеотида для G-белка (3) (рисунок 138(6)) (GEF-белок - Guanine nucleotide Exchange Factor (рисунок 79)).
Второй этап (рисунок 138(в,г,д)). G-белок диффундирует латерально вдоль мембраны и связывается с активированным гормоном мембранным GPCR-рецептором (рисунок 138(b)). Это событие стимулирует замену присоединённой к G-белку молекулы ГДФ на молекулу ГТФ (рисунок 138(г)), что в свою очередь вызывает резкое снижение сродства активированного гормоном мембранного рецептора (2) к комплексу G-белок+ГТФ (3) и к гормону (1) (рисунок 138(д)). G-белок, связанный с GTP, и гормон (1) уходят от рецептора (рисунок 138(e)).
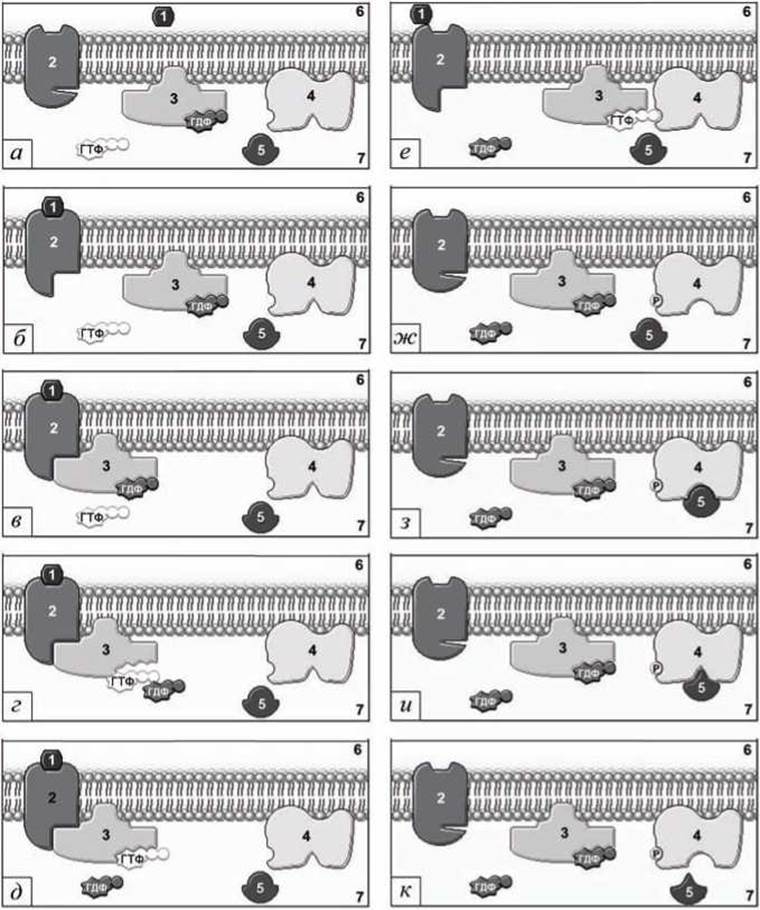
Рисунок 138 - Поэтапная схема мембранной рецепции: 1 - гормон (сигнальная молекула); 2 - рецепторный белок GPCR; 3 - G-белок; 4 - мембранный фермент; 5 - субстрат; 6 - межклеточное пространство; 7 - цитозоль клетки
Третий этап (рисунок 138(е,ж)). G-белок диффундирует вдоль мембраны и связывается с неактивным ферментом (4) (рисунок 138(e)). Крайняя фосфорильная группа ГТФ переносится на фермент. Фосфорилирование фермента вызывает изменение его конформации, формируется необходимая пространственная структура активного центра, и фермент становится каталитически активным (рисунок 138(ж)).
Четвёртый этап (рисунок 138(з,и,к)). Активированный фермент (4) связывает субстрат (5) (рисунок 138(з)) и катализирует изменение последнего (рисунок 138(и)). Преобразованный субстрат покидает фермент (рисунок 138(к)) и запускает внутриклеточные процессы, которые и являются ответом на внешний сигнал.
Если преобразованный субстрат активирует другие ферменты, то он и является вторичным месенджером.
Наиболее характерные свойства вторичного месенджера:
1. Вторичные месенджеры имеют относительно небольшую по сравнению с биополимерами молекулярную массу, поскольку молекула месенджера должна с высокой скоростью диффундировать в цитоплазме.
2. Вторичный месенджер обязан быстро (по сравнению со временем передачи сигнала) расщепляться, а в случае Са2+ - откачиваться из цитозоля. В противном случае сигнальная система останется во включённом состоянии и после того, как действие внешнего сигнала уже прекратилось.
Подобные ошибки могут оказаться в прямом смысле фатальными. Так, например, форболовые эфиры, которые представляют собой структурные аналоги диацилглицерола (рисунок 15(6)), но в отличие от него в организме не расщепляемые, способствуют развитию злокачественных опухолей. Это происходит потому, что форболовые эфиры вовлекаются в работу некоторых сигнальных систем, которые регулируют клеточное деление с помощью диацилглицерола как вторичного месенджера. Однако, имитируя действие ДАТ и обеспечивая передачу пролиферативного сигнала, они вовремя не расщепляются. В результате сигнальная система перестаёт чувствовать внешний сигнал и оказывается в перманентно включённом состоянии, а значит, пролиферация (деление) клеток перестаёт быть контролируемой.
Ниже мы рассмотрим некоторые примеры систем мембранной передачи сигналов.