Биологические мембраны - А. Н. Огурцов 2012
Электрогенез биомембран
Внутриклеточная сигнализация
цАМФ
Еще одним внутриклеточным месенджером является циклический аденозин монофосфат (цАМФ, сАМР) (рисунок 144(a)). Он используется во многих эукариотических клетках, например, в нейронах, обеспечивающих обоняние. Тело клетки такого нейрона находится в носу, а множественные дендриты располагаются в слизистой оболочке носа. В мембранах дендритов расположены белковые GPCR-рецепторы, специфические к "молекулам запахов" (scents). А аксон такого нейрона проникает в головной мозг.
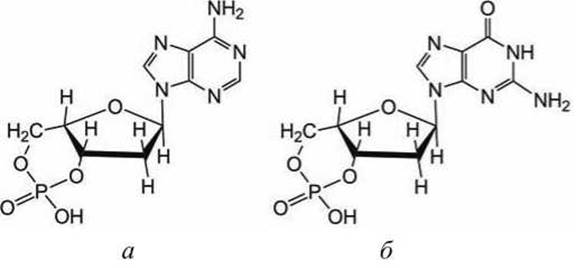
Рисунок 144 - Схемы циклических пуриновых нуклеозидмонофосфатов: а - цАМФ; б - цГМФ
Когда молекула пахучего вещества связывается с рецептором (рисунок 145), то рецептор, так же как и в случае чувствительности к АДФ, становится фактором замены нуклеотидов (GEF) для тримерного белка Gs, который функционально подобен тримерному белку Gq (рисунок 139).
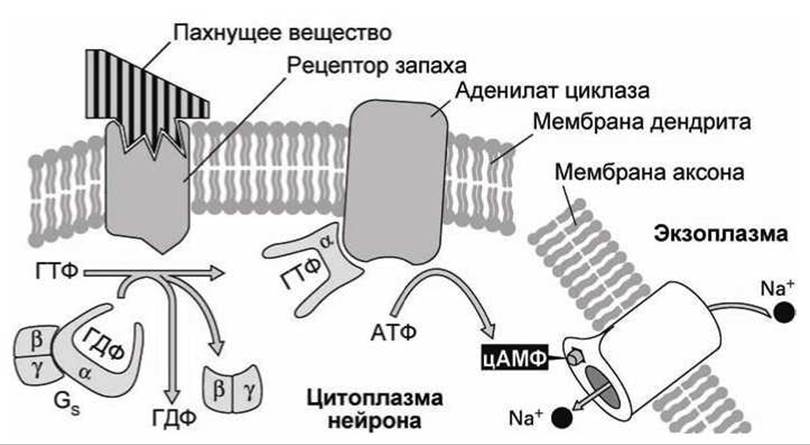
Рисунок 145 - Схема молекулярного механизма обоняния
Связанная с ГТФ субъединица а белка Gs в свою очередь активизирует мембранный фермент аденилат циклаза, который превращает АТФ в цАМФ. Затем цАМФ диффундирует в цитозоле нейрона к цАМФ-управляемым каналам в мембране аксона. Связывание с молекулой цАМФ открывает этот канал, что позволяет ионам калия и натрия перемещаться через мембрану. Электрохимический градиент стимулирует значительный поток ионов натрия внутрь клетки, что приводит к деполяризации мембраны, потенциалочувствительные натриевые каналы открываются, и развивается потенциал действия, который распространяется по аксону в головной мозг и сигнализирует о наличии пахнущего вещества. Белок Gs и фермент аденилат циклаза обнаружены во многих клетках, однако только в клетках нейронов, чувствительных к запаху, имеются рецепторы пахнущих веществ. Другие клетки используют внутриклеточный месенджер цАМФ для реагирования на те химические внеклеточные сигналы, для которых в мембранах клеток есть соответствующие рецепторы. После того, когда внешний химический сигнал исчезает, концентрация цАМФ снижается до исходного уровня. Фермент цАМФ-фосфодиестераза гидролизует цАМФ до АМФ, что инактивирует цАМФ-управляемые каналы и другие цАМФ-чувствительные белки.
С функционированием цАМФ-систем сигнализации напрямую связано патогенное действие возбудителя холеры кишечной бактерии Vibrio cholera. Токсин, который выделяет эта бактерия, является ферментом, который попадает в цитозоль клеток кишечного эпителия и присоединяет АДФ к Gs таким образом, что блокируется гидролиз ГТФ в ГДФ. Gs фиксируется в активном состоянии и непрерывно активизирует аденилат циклазу. В результате концентрация цАМФ в цитозоле растёт, все цАМФ-управляемые каналы открываются, из клеток уходят ионы и осмотически сопровождающие их молекулы воды. Происходит обезвоживание организма.