МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
Розділ 4. ГЕНЕТИКА БАКТЕРІЙ
ГЕНЕТИЧНИЙ АПАРАТ БАКТЕРІАЛЬНОЇ КЛІТИНИ
Генетика бактерій - це наука про спадковість і мінливість. Вона вивчає закономірності спадковості й мінливості мікробів та механізми передачі генетичної інформації від однієї клітини до іншої в межах одного або різних видів.
Біологічні властивості кожного організму визначаються його генотипом - сукупністю генів, локалізованих у нуклеоїді. Однак не вся інформація, яку має генотип, проявляється. У певних умовах бактерії, які утворилися з однієї батьківської клітини, можуть відрізнятися між собою за морфологічними ознаками, ферментативною активністю та ін. Сума реалізованих клітиною ознак у конкретних умовах середовища складає її фенотип.
Будова генетичного апарату прокаріотів тривалий час була дискусійним питанням, оскільки на відміну від вищих організмів вони не мають ні оформленого ядра, ні хромосом, аналогічних еукаріотам. У зв'язку з цим вважали, що бактерії в генетичному плані є анархічною групою біологічних об'єктів. Але, незважаючи на це, ще М. Бейєринк описав у бактерій зміни, які були стабільними, легко розпізнавалися та успадковувалися.
Роком народження генетики бактерій як науки вважається 1943, коли С. Лурія описав мутації стійкості щодо бактеріофагів. А за іншими даними, датою народження генетики мікроорганізмів є 1940 р., коли Г. Бідл і Е. Татум індукували мутації у Neurospora crassa і провели генетичний аналіз. На основі цих дослідів була сформульована концепція "один ген - один фермент», що поклало початок розвитку біохімічної генетики. У подальшому було встановлено, що ядерний апарат прокаріотів представлений ДНК, бактеріальне ядро функціонально тотожне ядру вищих організмів, але відрізняється деякими особливостями: ядерний апарат бактерій не має ядерної оболонки і ДНК перебуває в безпосередньому контакті з цитоплазмою; немає розподілу на хромосоми, а нитка ДНК - це аналог хромосоми вищих організмів, яка була названа бактеріальною хромосомою; у бактерій відсутній мітоз і мейоз.
Щоб підкреслити структурні відмінності генетичного апарату про- і еукаріотичних клітин, запропоновано в перших його називати нуклеоїдом, а у других - ядром. Нуклеоїд прокаріотів досить чітко відмежований від цитоплазми, компактно займає центральну частину клітини.
ДНК - макромолекула, при кислотному гідролізі якої відбувається її розщеплення на структурні елементи: дезоксирибозу, залишок фосфорної кислоти та азотисті основи (пуринові - аденін (А) і гуанін (G); піримідинові - цитозин (С) і тимін (Т)). У 1950 р. Є. Чаргаф установив ряд закономірностей щодо співвідношення азотистих основ:
- кількість аденіну в молекулі ДНК визначається в такій самій кількості, що й тиміну (А = Т), а вміст гуаніну дорівнює кількості цитозину (G = С);
- відношення G + C/A + T може варіювати в широких межах (у прокаріотів ці коливання перебувають у межах від 30 до 75 мол%, у той час як в еукаріотів - 35-40 мол%), але залишається постійним для конкретного виду;
- сума пуринових основ (А+G) дорівнює сумі піримідинових (C + T) : A + G = C + T.
У 1953 р. Д. Уотсон і Ф. Крик розшифрували структуру ДНК і запропонували модель подвійно закрученої спіралі (рис. 4.1).
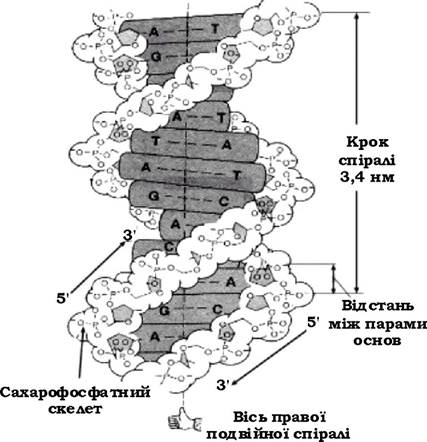
Рис. 4.1. Подвійна спіраль молекули ДНК
На один виток спіралі припадає близько 10 пар основ. Напрямок
ланцюгів протилежний (5' → 3' та 3' → 5'). Один мкм одноланцюгової ДНК відповідає відносній молекулярній масі приблизно 2 х 106 а.о.м. (атомна одиниця маси = 1,66057 х 10-27кг) або 3 000 пар основ. Таким чином, знаючи довжину хромосоми (у E. coli, наприклад, 1 400 мкм), можна визначити її молекулярну масу (2,9 х 109).
Величина геному у різних видів бактерій варіює від 0,8 х 106 до 8 х 106 пар основ. Для порівняння наводяться розміри геномів деяких еукаріотичних клітин: Neurospora crassa - 19 x 106; Aspergillus niger - 40 x 106; Homo sapiens - 2,9 х 106; Zea mays - 7 x 109пар основ.
Кількість геномів у клітинах різних видів бактерій може бути різною, вона залежить від умов культивування та фізіологічного стану культури. У періодичній культурі клітини E. coli, B. subtilis, що ростуть, мають від двох до восьми геномів, Azotobacter chroococcum - від 20 до 25, Desulfovibrio gigas - 10-15 геномів на одну клітину. При переході мікробної популяції у стадію стаціонарного росту у клітинах, як правило, визначається по одній хромосомі.
Існує думка, що нуклеоїд грампозитивних бактерій є більш компактною структурою, ніж грамнегативних, і займає відносно меншу частину клітини.
Молекула ДНК несе негативні заряди, оскільки кожен фосфатний залишок має у своєму складі іонізовану гідроксильну групу. В еукаріотів негативні заряди нейтралізуються утворенням комплексу ДНК з основними білками - гістонами. У більшості прокаріотів гістонів не знайдено, тому нейтралізація зарядів відбувається завдяки взаємодії ДНК із поліамінами (сперміном або спермідином), а також з іонами Mg2+. Щоправда, останнім часом у представників групи мікоплазм (Thermoplasma acidophilum), деяких ціанобактерій та архебактерій виявлено гістоноподібні білки, зв'язані з ДНК.
Спеціальними дослідженнями доведено, що хромосома прокаріотів - це високовпорядкована компактна структура. Така структура отримала назву "компактної хромосоми". Розрізняють дві форми хромосом: вільну і зв'язану. Вільна форма бактеріальної хромосоми має константу седиментації 1 300-2 000 S, а зв'язана з ЦПМ - 3 200-7 000 S.
Нуклеоїд вільний від елементів ЦПМ, містить в основному ДНК, РНК, а також гістоноподібні білки. РНК локалізується переважно в центральній частині нуклеоїду, що було доведено обробкою клітин РНКазою. При цьому центральна зона стає менш щільною, а константа седиментації знижується до 400-500 S. Така форма нуклеоїду має близько 140 петель, які відходять від щільної центральної зони (див. рис. 4.2). Нитка кожної петлі досягає 20 мкм завдовжки.
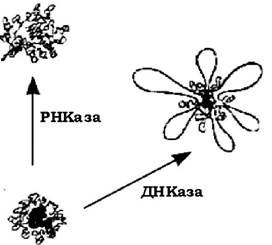
Рис. 4.2. Будова компактної хромосоми Escherichia coli
У зв'язаного фрагмента нуклеоїду визначаються зв'язки мембрани з петлями або центральною зоною. Обробка РНКазою знижує константу седиментації, що супроводжується збільшенням об'єму нуклеоїду. На цій підставі прийнято вважати, що компактність структури нуклеоїду забезпечується зв'язками типу РНК-РНК або РНК-ДНК. Збільшення об'єму нуклеоїду пояснюється втратою ДНК суперспіралізації, що супроводжується розширенням і розпрямленням петель. Зв'язок хромосоми з мембраною може здійснюватися двома шляхами: за рахунок зв'язку ДНК із специфічним білком мембрани або через рибосоми, зв'язані з мембраною. Кількість точок зв'язування досягає 15-20 і більше.
Модель структури активного нуклеоїду in vivo має враховувати високий ступінь зв'язку транскрипції та трансляції і можливість синтезу білкових молекул на рибосомах, зв'язаних з мембраною (рис. 4.3).
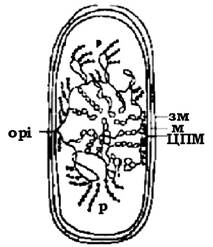
Рис. 4.3. Модель організації нуклеоїду Escherichia coli: орі - точка origin на хромосомі бактерії; р - рибосоми; зм - зовнішня мембрана; м - муреїн
Відповідно до такої моделі неактивна, суперспіралізована ДНК локалізована в центральній частині нуклеоїду, тоді як по його периферії розташовуються деспіралізовані петлі, які є матрицею синтезу іРНК. іРНК зв'язана з ДНК через РНК-полімеразу, а також із цитоплазматичними або мембранно-зв'язаними рибосомами. У більшості випадків бактеріальні хромосоми виявились кільцевими. Але в 1989 р. M. Фердоуз та A. Барбю, використовуючи пульс-форез, виявили нативну ДНК Borrelia burgdorfen, що мала лінійну форму. У подальшому лінійні структури бактеріальних хромосом були виявлені у актиноміцетів Streptomyces lividans, S. coelicolor, а також фітопатогенної бактерії Rhodococcus fascians. На обох кінцях таких структур розміщувались інвертовані повтори, до яких прикріплювались білкові молекули. Відомі випадки про наявність в одній бактеріальній клітині (Agrobacterium tumefaciens) двох хромосом, одна з яких мала кільцеву структуру, а друга лінійну. Не зрозуміло, як може суміщатись механізм реплікації кільцевої і лінійної структури хромосом в одній клітині.
Реплікація ДНК відбувається за напівконсервативним механізмом і в нормі завжди передує поділу клітини.
Оскільки один із ланцюгів ДНК містить послідовність нуклеотидів, яка є комплементарною іншому, тобто їх інформаційний зміст ідентичний, то будь-який з них може бути матрицею для нового ланцюга ДНК. У результаті утворюються два дуплекси ДНК, кожний з яких складається з одного ланцюга батьківської молекули ДНК і одного новосинтезованого (рис. 4.4).
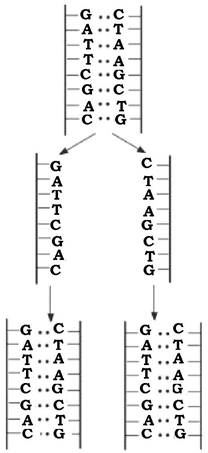
Рис. 4.4. Модель реплікації бактеріальної ДНК за напівконсервативним механізмом
Напівконсервативний механізм реплікації ДНК експериментально довели М. Мезельсон і Ф. Сталь у 1958 р. Процес реплікації ДНК здійснюється за участю багатьох білків, закодованих у послідовності нуклеотидів ДНК. Комплементарна будова на матричному ланцюзі ДНК здійснюється за допомогою ДНК-залежної ДНК-полімерази. За допомогою цього ферменту відбувається синтез ДНК на одноланцюговій матриці за наявності затравки - комплементарного фрагмента зростаючого ланцюга ДНК. ДНК-полімераза послідовно нарощує кінець затравки, приєднуючи до нього наступні нуклеотиди. Вибір чергового нуклеотиду для приєднання до затравки диктується матрицею і супроводжується гідролізом багатого енергією зв'язку і відщепленням пірофосфату. Останній розщеплюється пірофосфатазою, що робить реакцію незворотною. Синтез ДНК відбувається в напрямку 5' 3', при цьому 3'-ОН-група кінцевого нуклеотиду затравки атакує α-фосфат чергового дезоксирибонуклеотидтрифосфату, у результаті чого відщеплюється пірофосфат і дезоксирибонуклеотидмонофосфат зв'язується фосфодіефірним зв'язком зі зростаючим ланцюгом ДНК.
Головну роль в реплікації ДНК у E. coli виконує ДНК-полімераза ІІІ. Це великий мультимерний фермент, який має молекулярну масу 500 кД і складається із семи типів субодиниць: α, ε, θ, τ, γ, σ, β. Повна форма ферменту називається холоферментом. Вона забезпечує швидкість синтезу ДНК близько 1 000 нуклеотидів за секунду за оптимальних умов як in vitro, так і in vivo. Синтез ДНК розпочинається із синтезу РНК, яка служить РНК затравкою з декількох рибонуклеотидів і міститься на 5'-кінці. Затравка формується з допомогою ферменту - ДНК-праймази.
Батьківські молекули ДНК двоспіральні, тому перед реплікацією вони розділяються на матричні ланцюги. Цей процес здійснюється за рахунок хеліказ і SSB-білків. Хелікази - це ДНК-залежні АТФази, що використовують енергію гідролізу АТФ для розплетення подвійної спіралі ДНК. За рахунок гідролізу АТФ-хеліказа рухається по одному ланцюгу ДНК, розплітаючи перед собою подвійну спіраль. У результаті утворюється вилка реплікації. SSB-білки - це білки, що здатні зв'язуватись з однонитчастою ДНК. Вважається, що вони розправляють ДНК, витягують її і видаляють можливі елементи вторинної структури в комплементарних ділянках. Зв'язування SSB-білка з одноланцюговою ДНК активує ДНК-полімеразу і підвищує точність її роботи.
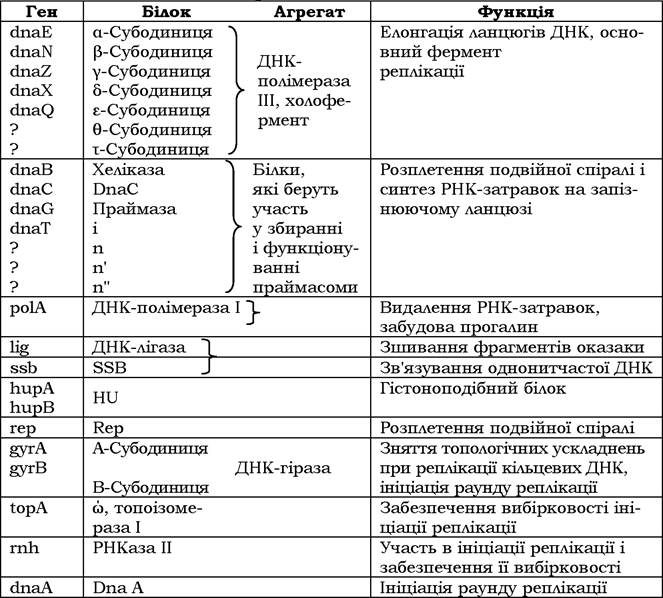
Відомо, що ланцюги ДНК антипаралельні, тому синтез їх на двох матричних ланцюгах відрізняється. Новосинтезована нитка, що утворюється безперервно, називається провідною, а інша - запізнюючою, на ній ДНК синтезується короткими фрагментами (фрагментами Оказакі ). Кожний фрагмент Оказакі (довжиною близько 1 000 нуклеотидів) має на 5'-кінці затравку із декількох рибонуклеотидів, утворених праймазою. Після синтезу фрагментів РНК-затравки видаляються, прогалини забудовуються ДНК-полімеразою, а потім фрагменти зшиваються в один неперервний ланцюг ДНК-лігазою. У реплікації ДНК E. coli беруть участь гени та їх продукти, наведені в табл. 4.1.
Реплікація ДНК супроводжується формуванням реплікативної вилки (рис. 4.5) за участю продуктів генів, наведених у табл. 4.1. Підраховано, що в реплікативній вилці одночасно працюють близько 20 білків, які здійснюють точний і складний процес реплікації.
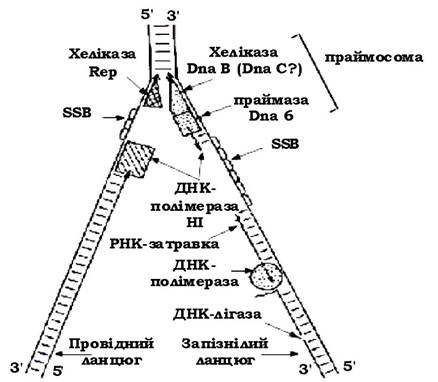
Рис. 4.5. Реплікація ДНК Escherichia coli в реплікативній вилці
Трансляція гену розпочинається із взаємодії рибосоми з мРНК, яка утворена лідируючою (5' - п'ятий атом рибози), білок-кодуючою і кінцевою (3' - третій атом рибози) частинами. Посадка здійснюється завдяки наявності в гені бактерій між промотором та ініціювальним кодоном сайта зв'язування рибосом. Він складається з 5-9 нуклеотидів, ядром яких є 5'AGGA3'. Посадка рибосом на мРНК здійснюється завдяки наявності комплементарних послідовностей нуклеотидів на 3' кінці 16S рРНК і в лідируючій частині мРНК.
Бактеріальна хромосома має окремі ділянки - гени (фрагменти молекули ДНК), які дискретно розташовані і несуть генетичну інформацію щодо всіх ознак, властивих клітині. Ген - головний фактор, який зумовлює спадкові властивості, тобто будь-яка спадкова ознака контролюється відповідними генами. Гени прокаріотів складаються із безперервно кодуючої послідовності нуклеотидів, тобто бактеріям властиве тісне зчеплення генів.
Гени, які несуть інформацію про синтез мікроорганізмами ферментів або структурних білків, називають структурними генами. Транскрипція структурних генів регулюється регуляторними генами.
Гени, або генетичні детермінанти, у бактерій організовані у відносно просту структуру, яка називається хромосомою. За своєю організацією вона відрізняється від хромосоми еукаріотів, проте генетичний матеріал (ДНК), який її складає, однаковий як для еукаріотів, так і для мікроорганізмів та ДНК-вмісних вірусів. Це і дає право називати структуру, яка містить генетичну інформацію бактерій, хромосомою.
Таблиця 4.1. Білки реплікації Escherichia coli та їхні гени
Гени, що контролюють синтез тієї чи іншої речовини, прийнято позначати маленькими буквами латинського алфавіту, які відповідають назві даної сполуки зі знаком "+" (his+ - гістидиновий ген, met+ - метіоніновий ген, leu+ - лейциновий ген та ін.). Гени, які контролюють резистентність до лікарських препаратів, фагів, отрут, позначають буквою r (лат. resistant - резистентний). Наприклад, резистентність до стрептоміцину записується як strr, а чутливість - strs. Фенотип бактерій позначають тими ж символами, що і генотип, але з великої букви.
Хромосома E. coli, наприклад, це кільцева молекула ДНК з молекулярною масою 2,9х106, яка містить дискретно розташовані гени. Циркулярну форму бактеріальної хромосоми було встановлено прямим методом за допомогою ауторадіографії і підтверджено в дослідах з генетичної рекомбінації. Ці методи дають змогу картувати розташування генів у бактеріальній хромосомі.
Генетична карта - це умовна схема хромосоми бактерій, в якій показана послідовність розташування окремих генів, їхня відносна протяжність і відстань між окремими генами, виражена в умовних одиницях рекомбінації (або в тисячах пар нуклеотидів - тпн) (рис. 4.6). Для побудови генетичних карт використовують різні способи перенесення генетичного матеріалу (кон'югація, трансформація, трансдукція). Висока роздільна здатність методів генетичного аналізу дозволила картувати хромосоми багатьох видів грампозитивних і грамнегативних бактерій та вивчати тонку будову і функції окремих генів.
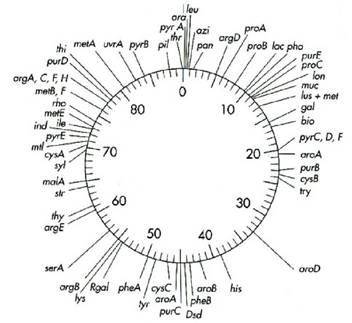
Рис. 4.6. Генетична карта Escherichia coli
Завдяки розробці різних і ефективних методик для генетичного і фізичного аналізу нуклеоїду, в E. coli картовано понад тисячу генів. Генетичні карти мікроорганізмів мають важливе не лише пізнавальне значення, вони можуть ефективно використовуватися в селективній роботі і генній інженерії.
Генетичний матеріал у прокаріотів може міститися не лише в хромосомі, а і в позахромосомних структурах - плазмідах, які можуть перебувати в цитоплазмі автономно або в інтегрованому з хромосомою стані. Плазміди - це молекули ДНК, які містяться в бактеріальних клітинах у вигляді закритих або відкритих кільцеподібних форм.