МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
Розділ 4. ГЕНЕТИКА БАКТЕРІЙ
ДИСОЦІАЦІЇ
П. Крюї, Дж. Аркрайт, а згодом й інші дослідники встановили своєрідну форму мінливості в бактерій, якою є S-R-дисоціація. Вона виникає спонтанно, її наслідком є утворення двох типів клітин, які формують різні форми колоній. При засіві на щільне середовище чистої культури утворюються колонії двох типів: гладенькі, S-форма (англ. smooth - гладенький) і шорсткі, R-форма (англ. rough - шорсткий). Процес дисоціації, тобто розщеплення бактеріальних клітин, які формують обидва типи колоній, зазвичай відбувається в одному напрямку: від S- до R-форми. Зворотний перехід R- у S-форму спостерігається рідше. Між цими двома типами колоній існують перехідні, нестійкі, частіше О-форми (рис. 4.7). Відмінності між S- і R-формами не обмежуються лише структурою колоній, вони супроводжуються зміною й інших ознак: фізіолого-біохімічних, патогенних властивостей бактерій, їх резистентності до дії фізичних і хімічних факторів середовища. Така мінливість отримала назву дисоціації (розщеплення). У процесі дисоціацій можуть утворюватися й інші форми колоній: карликові (D-dward), G-колонії (gonidial), які формуються на поверхні або біля країв нормальних колоній.
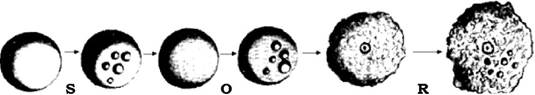
Рис. 4.7. Зміна форми колоній у процесі дисоціації:
S - гладенька; О - мутна; R — шорстка
Дисоціації розглядають як одну із форм внутрішньовидової мінливості, при якій в однорідній популяції розвиваються різні за біологічними властивостями та ознаками мікроорганізми, що утворюють декілька типів колоній (табл. 4.2). Проте найбільш чітко змінені ознаки та властивості бактерій спостерігаються у крайніх - S- і R-типах колоній.
Таблиця 4.2. Ознаки та властивості S- і R-форм колоній
Ознака |
S-форма |
R-форма |
Характер росту |
Гомогенний ріст при культивуванні в рідкому поживному середовищі. На твердих поживних середовищах ріст рівномірний, колонії вологі, блискучі, опуклі, гладенькі |
Аглютинативний ріст у вигляді дрібнозернистого осаду. Ріст на твердих середовищах сухий, щільний, колонії шорсткі, плоскі |
Морфологія |
Клітини нормальної морфології. У капсульованих видів наявні капсули, у рухомих - джгутики |
Клітини зменшені в розмірах, кокоподібні. У капсульних форм капсули відсутні, рухомі клітини можуть не мати джгутиків |
Патогенність |
У патогенних видів проявляється висока вірулентність |
Слабо або зовсім не вірулентні |
Антигенні властивості |
Повноцінний антиген. Містять специфічні антигени. В імунній сироватці дають осад у вигляді пластівців |
Неповноцінні антигени. Часто містять неспецифічні антигени. В імунній сироватці формується дрібнозернистий осад |
Біохімічні властивості |
Біохімічно більш активні |
Менш активні |
Чутливість до: - бактеріофага; - фагоцитозу; - УФ-променів |
Чутливі; фагоцитуються слабо; чутливі |
Менш чутливі; легко фагоцитуються; стійкі |
Виділення з організму |
У гострий період захворювання |
При хронічному перебігу інфекційного процесу або при бацилоносійстві |
Біологічне значення S-R-дисоціації полягає в набутті бактеріями певних селективних переваг, які забезпечують їхнє існування в організмі людини або в навколишньому середовищі. До таких переваг належать висока стійкість S-форм до фагоцитозу макрофагами та бактерицидної дії сироватки крові. R-форми проявляють більш високу стійкість до дії факторів довкілля.
Поряд з цим S-R-дисоціація в багатьох випадках ускладнює бактеріологічну діагностику деяких інфекційних захворювань, зокрема дизентерії Зонне, ешерихіозу, викликаного E.coli 0124 та ін.
Мутації (лат. mutatio - зміна) - це зміна кількості або порядку локалізації нуклеотидів у генетичному матеріалі бактеріальної клітини, які супроводжуються зміною ознак або властивостей, що передаються з покоління в покоління.
Усе різноманіття мутацій поділяється на певні групи. За походженням розрізняють спонтанні й індуковані мутації. Спонтанними називають мутації, поява яких викликана невідомим фактором, без втручання дослідника. Такі мутації характеризуються випадковістю (мають ненаправлений характер), низькою частотою (1 х 10-10-1 х 10-5) та наявністю лаг-періоду (уповільнення їхнього фенотипового прояву), тобто вони проявляються після декількох циклів клітинного поділу.
Спонтанні мутації виникають у результаті помилок у репарації (відновленні), реплікації і рекомбінації ДНК під впливом природного фону опромінення, ультрафіолетових (УФ) променів або дії хімічних речовин. Ці мутації є основним джерелом природної мінливості мікроорганізмів і лежать в основі їхньої еволюції. Однією з причин виникнення спонтанних мутацій вважаються помилки у функціонуванні ДНК-полімерази, які виникають під час реплікації ДНК, коли цей фермент уводить в новосинтезовану нитку замість однієї азотистої основи іншу некомплементарну.
Відома наявність у геномі бактерій так званих генів-мутаторів, мутація в яких більш ніж у 100 разів підвищує кількість спонтанних мутацій.
Індуковані мутації виникають під дією певного мутагенного фактора. Мутагенні фактори можуть мати хімічну, фізичну або біологічну природу.
Хімічні мутагени - це речовини, які викликають хімічні перетворення, що супроводжуються змінами у структурі ДНК. До них належать: інгібітори синтезу попередників ДНК; аналоги азотистих основ; алкілувальні сполуки; похідні акридину; речовини, які змінюють структуру азотистих основ.
Інгібітори синтезу попередників нуклеїнових кислот - це речовини, які перешкоджають утворенню азотистих основ, наприклад: азасерин, бензимідазол, теобромін, тетраметилсечова кислота, етилуретан та ін. Вважається, що ці речовини інгібують синтез азотистих основ.
Аналоги азотистих основ - це речовини, які здатні замінювати азотисті основи в молекулі ДНК: 5-бромурацил, 5-бромурацилдезок- сирибоза та ін. Так, перший із них займає місце тиміну, а переходячи
в енольну форму, набуває здатності спарюватися з гуаніном, тобто утворювати незаконну пару основ. 2-амінопурин є аналогом аденіну. Замість тиміну він спаровується з цитозином, у результаті чого відбуваються зміни пар основ АТ → ГЦ.
Алкілувальні сполуки - це речовини, у присутності яких до певної частини молекули ДНК приєднуються залишки вуглеводнів (алкілів) - метильний, етильний та ін. До таких мутагенів належать: етилетансульфонат, метилметансульфонат, етилметансульфонатдиметилсульфат, етиленамін та ін. Алкілувальні сполуки взаємодіють із фосфатною групою, утворюючи нестійкі сполуки алкілу з фосфатом. Крім того, алкільна група може заміщувати водень і приєднуватися через кисень, азот або сірку. Найчастіше в ДНК алкілується гуанін в 7-му положенні.
Багато нітрозосполук (нітрозогуанідин, нітрозосечовина та ін.) мають високу мутагенну активність, тому вони отримали назву супер- мутагенів.
Похідні акридину вводяться між сусідніми основами ланцюга ДНК, збільшуючи відстань між ними. При реплікації це часто викликає втрату або включення додаткового нуклеотиду, що супроводжується порушенням порядку зчитування інформації у процесі синтезу білка. Акридинові барвники можуть індукувати випадання однієї або декількох пар азотистих основ.
Речовини, які змінюють структуру азотистих основ, наприклад азотиста кислота, викликають реакцію окиснювального дезамінування. При цьому аденін перетворюється в гіпоксантин, який спаровується не з тиміном, як аденін, а з цитозином. При окиснювальному дезамінуванні цитозину утворюється урацил, який спаровується не з гуаніном, а з аденіном.
Фізичні фактори - це насамперед короткохвильові випромінювання (ультрафіолетові та іонізуючі випромінювання - рентгенівські, Ύ-про- мені, β-частинки, протони, нейтрони, α-частинки) і дія температури.
Іонізуючі промені проникають у клітину і викликають іонізацію атомів і молекул речовин. Відбувається відрив електронів від молекул однієї речовини (вони перетворюються в позитивно заряджені частинки), а електрони, які відірвалися, приєднуються до молекул іншої речовини, перетворюючи їх у негативно заряджені частинки. Одна і та сама речовина, наприклад вода, може утворювати пару іонів. Під впливом опромінення від молекули води відривається електрон і вода набуває позитивного заряду. Інша молекула води приєднує електрон і набуває негативного заряду:
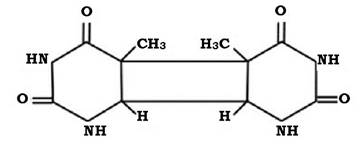
Іонізовані молекули нестійкі, вони розкладаються з подальшою рекомбінацією радикалів і утворенням стійких молекул - H2O, H2 і Н2О2. Пероксид водню під впливом каталази розпадається на О2 і Н2О. Під впливом пероксиду водню й активного кисню можуть утворюватися перекисні органічні сполуки, які вступають у реакції з нуклеїновими кислотами, змінюючи їхню структуру.
Ультрафіолетові промені викликають пошкодження, пов'язані, головним чином, з утворенням димерів тиміну. На такий тип пошкодження, як піримідинові димери, припадає ~90 % мутацій, індукованих УФ-променями.
Подібні факти свідчать про існування деякої внутришньогенної специфічності мутагенів, яка полягає в тому, що деякі мутагени найчастіше вражають певні специфічні ділянки в одному і тому ж гені або викликають переважно один і той же тип пошкодження структури ДНК.
Дія температури є слабким мутагенним фактором, але її підвищення на кожні 100С збільшує частоту генних мутацій у декілька разів, підсилює дію інших фізичних та хімічних мутагенів.
Біологічні мутагени - це віруси бактерій, інсерційні (IS-елементи), транспозони (Tn-елементи), що є мігруючими генетичними елементами. ДНК вірусів здатна інтегруватися в геном бактеріальної клітини, що супроводжується дестабілізацією сусідніх генів. Мігруючі генетичні елементи виявлені в ДНК усіх бактерій. Ці фрагменти ДНК здатні до внутрішньо- і міжхромосомних переміщень, викликаючи індукцію таких порушень, як делеції, транслокації, реверсії, злиття і дроблення репліконів. Ці мутації можуть утворюватися на рівні генів, хромосом і плазмід. Кінці у IS-елементів і транспозонів фланковані прямими або інвертованими (ACTG...TGAC) повторами. Така структура отримала назву транспозонної (англ. transpose - переміщувати). IS-елементи та транспозони можуть існувати і відтворюватися лише в хромосомах, плазмідах і вірусах.
Як індуковані, так і спонтанні мутації за характером перебудов, які відбуваються в ДНК, поділяються на точкові і хромосомні.
Точкові мутації, при яких відбуваються хімічні зміни лише одного нуклеотиду, називають генними. Точкові мутації поділяють на декілька груп, які відрізняються за характером змін у ДНК, зумовлених мутагенним фактором. При мутаціях, які називаються транзиціями (прості заміни), пурин в одному з ланцюгів ДНК заміщується іншим пурином (аденін![]() гуанін) або піримідин у комплементарному ланцюзі - іншим піримідином (тимін
гуанін) або піримідин у комплементарному ланцюзі - іншим піримідином (тимін![]() цитозин). Зміни, при яких відбувається заміна пурину піримідином, називають трансверсіями (складні заміни). До точкових мутацій відносять також вставку додаткового нуклеотиду. Такі мутації становлять групу так званих мутацій із зсувом рамки, при яких відбувається зміщення читання послідовності нуклеотидів у ДНК.
цитозин). Зміни, при яких відбувається заміна пурину піримідином, називають трансверсіями (складні заміни). До точкових мутацій відносять також вставку додаткового нуклеотиду. Такі мутації становлять групу так званих мутацій із зсувом рамки, при яких відбувається зміщення читання послідовності нуклеотидів у ДНК.
Хромосомні мутації зв'язані з перебудовами в окремих фрагментах ДНК (рис. 4.8). Вони проявляються в результаті випадіння меншої або більшої кількості нуклеотидів (делеції), повороту певної ділянки ДНК на 1800 (інверсії) або повторення якого-небудь фрагмента ДНК (дуплікації).
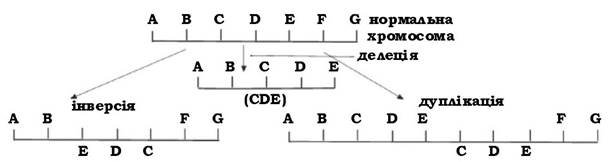
Рис. 4.8. Типи хромосомних мутацій
За напрямком зміни ознаки мутації бувають прямі і зворотні. При прямих мутаціях відбуваються зміни в генах дикого типу бактерій (наприклад, поява ауксотрофії у прототрофних штамів). Зворотні мутації - це зміни від мутантного типу до дикого. При цьому можуть бути істинні зворотні мутації, при яких має місце відновлення генотипу або супресії - відновлення фенотипу, тобто виникає вторинна мутація, яка пригнічує прояв першої.
Мутанти, які втрачають здатність до синтезу деяких речовин (амінокислот, пуринів, піримідинів, вітамінів та ін.), необхідних для їх розвитку, називають ауксотрофними. Ауксотрофні мутанти знайшли своє практичне використання. Так, у дикого штаму Corynebacterium glutamicum, лізин синтезується за звичайною схемою з утворенням деяких інших продуктів - гомосерину, треоніну, ізолейцину тощо, тобто вихідний субстрат (аспарагінова кислота) витрачається на синтез не лише лізину, а й інших речовин, що знижує вихід цільового продукту (лізину). При блокуванні синтезу гомосериндегідрогенази весь напівальдегід аспарагінової кислоти трансформується до лізину (рис. 4.9).
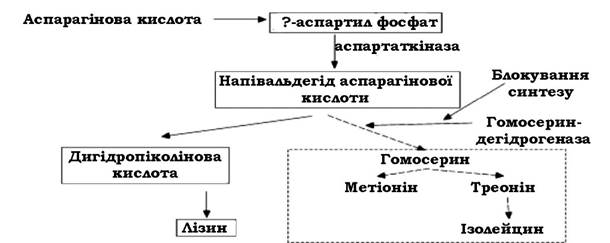
Рис. 4.9. Схема синтезу лізину ауксотрофними мутантами
Мутація - це стабільна зміна генетичного матеріалу, яка реалізується тими ж каналами, що і будь-яка інша інформація. Але доля мутацій може бути різною. Деякі з них не впливають на ознаки організму, залишаються "мовчазними". Якщо мутація призводить до зміни в активному центрі ферменту, то це відразу ж відбивається на його функціях. Діапазон таких змін може бути різний: від незначного зниження активності ферменту до повної її втрати. В останньому випадку такі зміни можуть бути летальними для клітини, в якій виникла мутація.