МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
Розділ 5. МЕТАБОЛІЗМ МІКРООРГАНІЗМІВ
МЕХАНІЗМИ АСИМІЛЯЦІЇ ВУГЛЕЦЮ МІКРООРГАНІЗМАМИ
Джерелами вуглецю для мікроорганізмів можуть бути органічні та неорганічні речовини (вуглекислий газ, карбонати).
Асиміляція неорганічного вуглецю. Вуглекислий газ - це найпоширеніша форма вуглецю на Землі. Багато мікроорганізмів, що називаються автотрофами, мають здатність включати СО2 до вуглецю клітин. Це явище отримало назву фіксації СО2.
Фіксація вуглекислого газу. У мікроорганізмів існую чотири шляхи фіксації вуглекислого газу:
1. Цикл Кальвіна (відновлювальний пентозофосфатний шлях або рибулозодифосфатний шлях, RuBP);
2. Відновлювальний цикл трикарбонових кислот (цикл Еванса - Буханана - Арнона, rTCA);
3. Відновлювальний шлях ацетил-КоА (відновлювальний шлях Люнг- даля - Вуда, rACA);
4. 3-Гідроксипропіонатний цикл.
Цикл Кальвіна (рис. 5.30) - найбільш важливий шлях біосинтезу на Землі. Його використовує більшість фотосинтезуючих організмів: рослини, ціанобактерії, пурпурні та зелені бактерії. Цикл Кальвіна складається з трьох фаз:
1. Фіксування вуглекислого газу. СО2 зв'язується з рибулозо-1,5- дифосфатом - сполукою, що містить п'ять атомів вуглецю. У результаті утворюється дві молекули 3-фосфогліцеринової кислоти. Одна з двох карбоксильних груп цієї сполуки походить із СО2. Реакцію каталізує рибулозо-1,5-дифосфаткарбоксилаза (RuBisCo) - найбільш унікальний ключовий фермент цього циклу.
Рибулозо-1,5-дифосфат + СО2 + Н2О → 3-Фосфогліцеринова кислота (2 молекули)
2. Відновлення карбоксильних груп фосфогліцеринової кислоти до альдегіду. Спочатку активується карбоксильна група 3-фосфогліцеринової кислоти шляхом фосфорилювання з утворенням 1,3-дифосфогліцеринової кислоти, яка потім відновлюється до 3-фосфогліцеринового альдегіду. На цій стадії витрачається практично вся енергія та відновник, необхідні для фіксації СО2. Частково 3-фосфогліцериновий альдегід перетворюється у фруктозо-6-фосфат і йде на потреби біосинтезу.
3-Фосфогліцеринова кислота + АТФ → 1,3-Дифосфогліцеринова кислота + АДФ
1,3-Дифосфогліцеринова кислота + НАДФН2 → 3-Фосфогліцериновий альдегід + НАДФ+ + Фн
3. Регенерація акцептора СО2. Решта реакцій циклу призводять до відтворення рибулозо-1,5-дифосфату. Розпочинаються вони синтезом фруктозо-6-фосфату з двох молекул тріозофосфатів. Потім проходять реакції перебудови вуглецевого скелету з утворенням пентозо-5-фосфатів, що нагадують обернені (відновлювальні) реакції пентозо- фосфатного шляху. Регенерація акцептора СО2 завершується фосфорилюванням рибозо-5-фосфату до рибулозо-1,5-дифосфату із витратою АТФ. Процес каталізується другим специфічним ключовим ферментом циклу Кальвіна - фосфорибулокіназою.
Енергетичний баланс циклу Кальвіна. Для утворення однієї додаткової молекули фруктозо-6-фосфату, яка буде виведена із циклу і використана для біосинтезу, має карбоксилюватися шість молекул рибулозо- 1,5-дифосфату, тобто зафіксуватися 6СО2. Шість молекул рибулозо-1,5-дифосфату після карбоксилювання перетворяться у 12 молекул 3- фосфогліцеринової кислоти, потім у 12 молекул 1,3-дифосфогліцеринової кислоти, які згодом відновляться до 12 молекул 3-фосфогліцеринового альдегіду. На такі перетворення клітина витратить 12 АТФ та 12 НАДФН2. Наступні 6 АТФ будуть використані при регенерації акцептора СО2 на фосфорилювання рибозо-5-фосфату до рибулозо-1,5-дифосфату. Таким чином, синтез однієї молекули фруктози "коштує" клітині 18 АТФ та 12 НАДФН2. Загальне стехіометричне рівняння фіксації СО2 через цикл Кальвіна має такий вигляд:
6СО2 + 12 НАДФН2 + 18 АТФ → С6Н12О6 + 12 НАДФ+ + 18 АДФ + 18 Фн
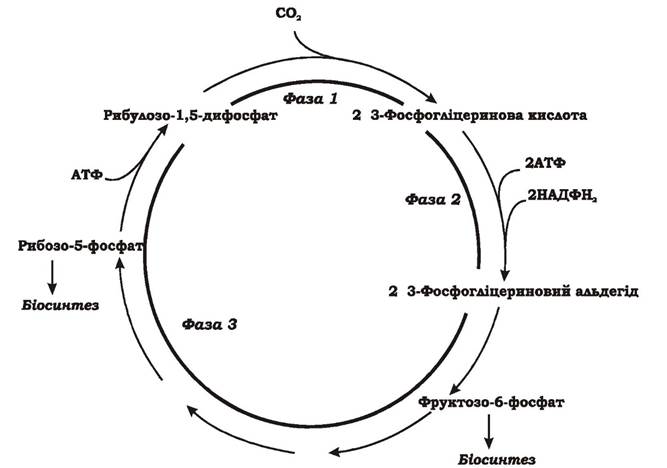
Рис. 5.30. Цикл Кальвіна
(відновлювальний пентозофосфатний шлях або рибулозодифосфатний шлях)
Відновлювальний цикл трикарбонових кислот був описаний Евансом, Бухананом та Арноном у 1966 р. у зеленої сіркової бактерії Chlorobium thiosulfatophilum. Пізніше його було також виявлено в деяких сульфатредукторів і термофільних водневих бактерій Hydrogenobacter. Відновлювальний ЦТК - це модифікований цикл Кребса, який функціонує у зворотному, відновлювальному напрямку (рис. 5.31).
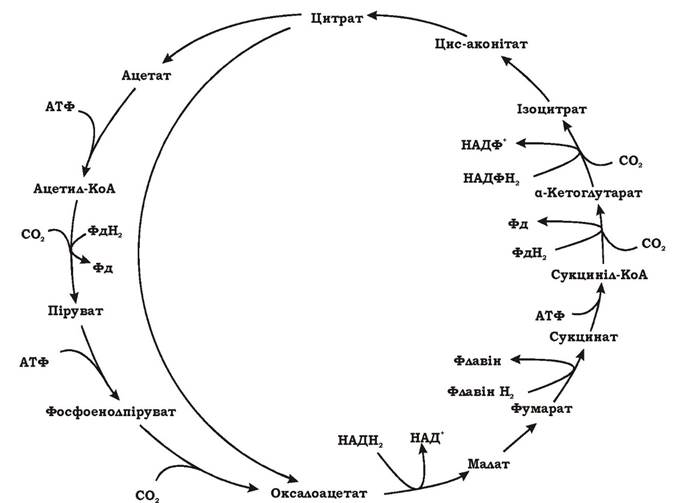
Рис. 5.31. Відновлювальний цикл трикарбонових кислот (синоніми: цикл Еванса - Буханана - Арнона, відновлювальний ЦТК)
Розрізняють повний і короткий відновлювальні ЦТК. У результаті одного оберту повного циклу фіксується чотири молекули СО2 та синтезується ЩОК. Оскільки ЩОК сама по собі є проміжним продуктом циклу, то після одного обертання одна молекула ЩОК регенерується, а друга синтезується з чотирьох фіксованих і відновлених СО2. За короткий цикл дві зафіксовані молекули СО2 утворюють одну молекулу ацетату.
Як видно з рисунка, повний і короткий цикли мають однакову послідовність реакцій від ЩОК до цитрату. Тобто, починаючи з ЩОК, повне обертання короткого відновлювального ЦТК призводить до регенерації ЩОК і синтезу ацетату з СО2. У підсумку саме короткий відновлювальний ЦТК, який утворює ацетил-КоА з 2СО2, є оберненим до циклу Кребса. У повному відновлювальному ЦТК мають місце чотири реакції фіксації та відновлення СО2:
1. Ацетил-КоА + СО2 + Фередоксинвідн → Піруват + КоА + Фередоксинокис
2. Сукциніл-КоА + СО2 + Фередоксинвідн → α-Кетоглутарат + КоА + Фередоксинокис
3. α-Кетоглутарат + СО2 + НАДФН2 → Ізоцитрат + НАДФ+
4. Фосфоенолпіруват + СО2 → ЩОК + Фн
Реакції (2) та (3) існують як у звичайному циклі Кребса, так і у відновлювальному ЦТК, а реакції (1) та (4) - лише у відновлювальному ЦТК. Реакції (і) і (2) відновлюються за рахунок фередоксинів, а не НАДФН2. Асиміляція СО2 супроводжується з витратами енергії. Відкриття циклу Еванса - Буханана - Арнона підтвердило еволюційний зв'язок між гетеротрофними та фотоавтотрофними мікроорганізмами.
Відновлювальний шлях ацетил-КоА зустрічається в метаболізмі автотрофних сульфатредукторів, гомоацетогенів та метаногенів. На відміну від решти шляхів асиміляції СО2, він не є циклічним (рис. 5.32). Одна молекула СО2 зв'язується спеціальним кофактором - тетрагідрофолієвою кислотою і відновлюється до метильної групи. Друга молекула СО2відновлюється до карбонільної (С=О) групи за допомогою ферменту карбо- нмонооксиддегідрогенази. Метильна та карбонільна групи об'єднуються в ацетил-КоА, який використовується в подальших процесах біосинтезу. Дві молекули СО2 відновлюється за рахунок 4Н2.
Загальне рівняння процесу: 4Н2 + 2СО2 = Ацетат + 2Н2О
Шлях ацетил-КоА може проходити і у зворотному, окисному напрямку. Окисний шлях ацетил-КоА використовують гетеротрофні сульфатредуктори для розщеплення ацетату (рис. 6.26).
3-Гідроксипропіонатний цикл фіксації СО2 (рис. 5.33) було виявлено в зелених несіркових фотосинтезуючих бактерій роду Chloroflexus та деяких археїв.
У результаті повного обертання цього циклу фіксується 2СО2 та синтезується гліоксилат (СНОСООН). Перша молекула СО2 акцептується ацетил-КоА з утворенням малоніл-КоА, а друга молекула СО2, з'єднуючись з пропіоніл-КоА, утворює метилмалоніл-КоА. Кінцевий проміжний продукт циклу - маліл-КоА розщеплюється на ацетил-КоА та гліоксилат. Ацетил- КоА знову фіксує СО2 на наступному оберті, а гліоксилат використовується для біосинтезу. Процес можна описати таким рівнянням:
2НСО3- + 2 НАДФН2 + 3Н+ + 3 АТФ → СНО-СОО- + 2 НАДФ+ + 3 АДФ + 3 Фн
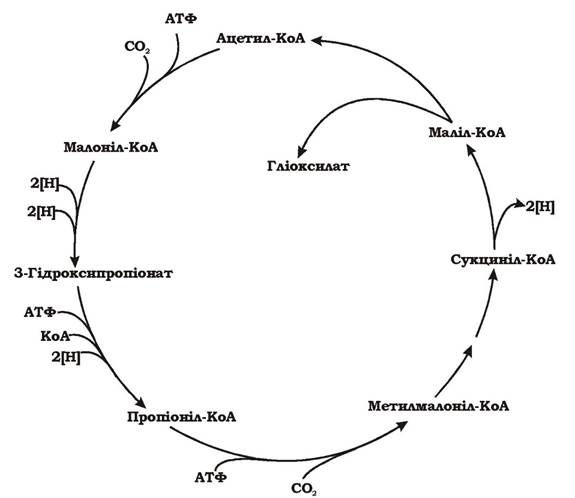
Рис. 5.32. Відновлювальний шлях ацетил-КоА (синонім: відновлювальний шлях Люнгдаля - Вуда): THF - теграгідрсфолієва кислота
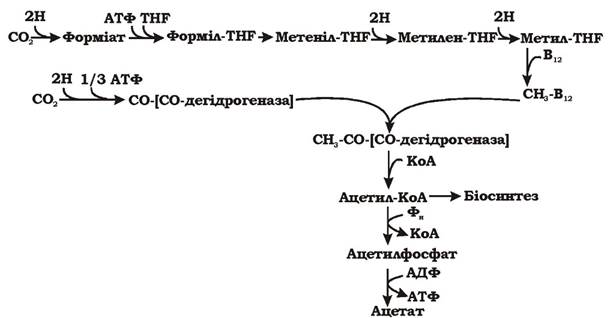
Рис. 5.33. 3-Гідроксипропіонатний цикл
Донори електронів та природа багатьох ферментів цього циклу не вивчені.
Асиміляція формальдегіду. У метилотрофних мікроорганізмів існує три способи асиміляції формальдегіду (СН2О):
1. Сериновий шлях.
2. Рибулозомонофосфатний цикл (гексулозофосфатний шлях, RuMP).
3. Дигідроксіацетоновий шлях.
Перші два шляхи характерні для метилотрофних бактерій, останній - лише для метилотрофних дріжджів.
Сериновий шлях - це циклічний процес, кінцевим продуктом якого є ацетил-КоА, що синтезується з однієї молекули формальдегіду та однієї молекули СО2 (рис. 5.34).
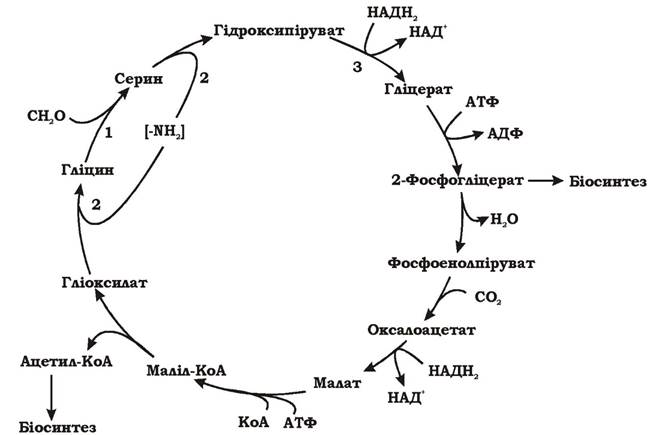
Рис. 5.34. Сериновий шлях асиміляції формальдегіду
У результаті першої реакції цього циклу при взаємодії формальдегіду (С1) з гліцином (С2) утворюється серин (С3). Реакція каталізується гідроксиметилтрансферазою, яка містить кофактор тетрагідрофолієву кислоту (THF). Процес іде через проміжні продукти, зв'язані з THF. При приєднанні формальдегіду до кофактора гідроксиметилтрансфе- рази утворюється 5,10-метилентетрагідрофолієва кислота (метилен-
THF). Внаслідок реакції (1) (рис. 5.34, 1) формальдегід переноситься з метилен-THF на гліцин і утворюється серин.
У наступних реакціях (рис. 5.34, 2) відбувається перенесення аміногрупи з серину на гліоксилат, у результаті чого з серину утворюється гідроксипіруват, а з гліоксилату - гліцин. Гліцин взаємодіє з наступною молекулою формальдегіду, а гідроксипіруват перетворюється далі (рис. 5.34, 3).
Гідроксипіруват відновлюється до гліцеринової кислоти, гліцеринова кислота фосфорилюється до 2-фосфогліцерату з використанням 1 АТФ. На цьому етапі сериновий шлях розщеплюється: частина 2-фосфогліцеринової кислоти перетворюється у 3-фосфогліцеринову кислоту, що йде на біосинтез, а решта перетворюється у фосфоенолпіруват (ФЕП). ФЕП (С3) карбоксилюється СО2 до оксалоацетату (С4), який відновлюється до малату і з використанням АТФ перетворюється в маліл-КоА. За допомогою маліл-КоА-ліази маліл-КоА розщеплюється на гліоксилат (С2-сполука), який бере участь у циклі, та на ацетил-КоА (С2-сполука), що йде на біосинтез.
Рибулозомонофосфатний цикл (рис. 5.35) - це циклічний процес асиміляції формальдегіду, який складається з трьох стадій:
1. Фіксація. Акцептором формальдегіду є рибулозо-5-фосфат. Формальдегід та рибулозо-5-фосфат конденсуються за допомогою гексулозофосфатсинтетази в гексулозо-6-фосфат, який потім перетворюється у фруктозо-6-фосфат під дією ферменту гексулозофосфатізомерази. Тільки гексулозофосфатсинтетаза та гексулозофосфатізомераза є унікальними ферментами для цього шляху. На інших стадіях реакції каталізують ферменти шляху Ентнера - Дудорова.
2. Розщеплення. Фруктозо-6-фосфат розщеплюється до С3-сполук двома способами:
а) фруктозо-6-фосфат фосфорилюється до фруктозо-1,6-дифосфату, а потім розщеплюється на діоксіацетонфосфат та фосфогліцериновий альдегід;
б)фруктозо-6-фосфат спочатку ізомеризується до глюкозо-6-фосфату, який перетворюється у Б-глюконо-1,5-лактон-6-фосфат, а згодом - у 6-фосфоглюконат, і нарешті - у 2-кето-3-дезокси-6-фосфоглюконову кислоту (КДФГ). КДФГ розщеплюється на 3-фосфогліцериновий альдегід та піруват за допомогою КДФГ-альдолази.
Одна з фосфотріоз (діоксіацетонфостат чи піруват) подається на біосинтез.
3. Перегрупування. Остання стадія процесу - це перегрупування фосфотріоз, при якому відбувається регенерація акцептора формальдегіду - рибулозо-5-фосфату. Воно може здійснюватися кількома шляхами, але в результаті дві молекули ксилулозо-5-фосфату та рибозо-5-фосфат перетворюються у три молекули рибулозо-5-фосфату. Цикл замикається.
Отже, в результаті рибулозомонофосфатного циклу з трьох молекул формальдегіду утворюються тріоза (діоксіацетонфостат чи піруват), яка подається на біосинтез.
Загальне рівняння процесу:
ЗСН2О + 2 АТФ → Фосфотріоза + 2 АДФ + Фн
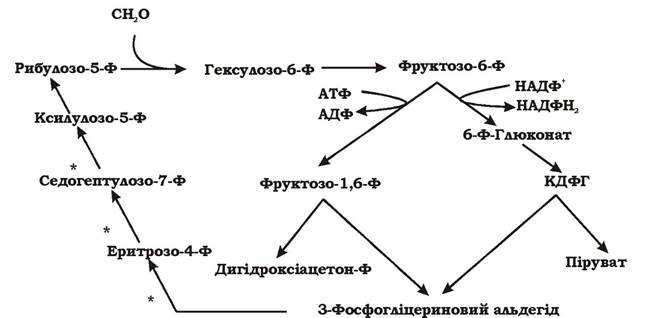
Рис. 5.35. Схема рибулозомонофосфатного циклу асиміляції формальдегіду (синонім: гексулозофосфатний шлях, RuMP)
* - позначено реакції, які можуть відбутися лише при включенні у процес не менше трьох молекул формальдегіду
Асиміляція С2-субстратів мікроорганізмами. При рості на середовищах з ацетатом, як єдиному джерелі вуглецю, аеробні мікроорганізми асимілюють ацетил-КоА через гліоксилатний цикл.
Гліоксилатний цикл - це послідовність реакцій, яка перетворює дві ацетильні одиниці в одну С3-сполуку з декарбоксилюванням четвертого атома вуглецю: 2С2 → 1С3 + 1С1 (рис. 5.36).
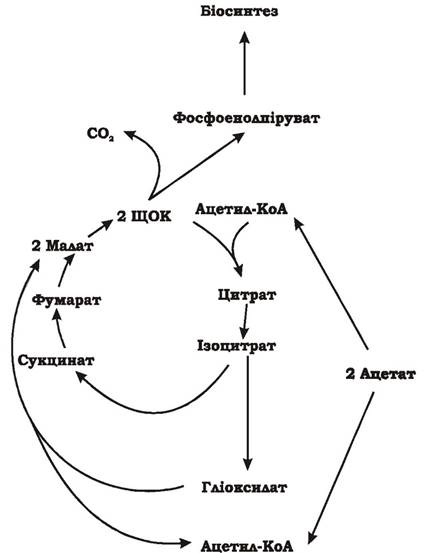
Рис. 5.36. Гліоксилатний цикл асиміляції ацетил-КоА мікроорганізмами
На першому етапі цього циклу ацетил-КоА з'єднується зі ЩОК і перетворюється в цитрат, а згодом - в ізоцитрат, який під дією ізоцитратліази розщеплюється на два проміжних продукти: гліоксилат і сукцинат.
Ізоцитрат → Гліоксилат + Сукцинат
На другому етапі синтезується дві молекули малату. Одна молекула утворюється безпосередньо при перетворенні сукцинату через фумарат, друга - за рахунок реакції конденсації гліоксилату, що утворився на першому етапі, та другої молекули ацетил-КоА. Ця реакція каталізується малатсинтетазою:
Гліоксилат + Ацетил-КоА + Н2О → Малат + КоА-SH
На третьому етапі з двох молекул малату синтезується дві молекули ЩОК, одна з яких подається на біосинтез, а друга - функціонує у гліокилатному циклі. Для включення в біосинтез ЩОК декарбоксилюється до пірувату чи фосфоенолпірувату.
Глюконеогенез - це процес синтезу глюкози з пірувату (рис. 5.37). Він подібний до гліколізу, де глюкоза окиснюється до пірувату, але реакції проходять у зворотному (відновлювальному) напрямку.
Глюконеогенез і гліколіз мають також ряд суттєвих розбіжностей. На відміну від гліколізу, де при окисненні глюкози утворюється 2 АТФ, при синтезі глюкози шляхом глюконеогенезу витрачається 6АТФ.
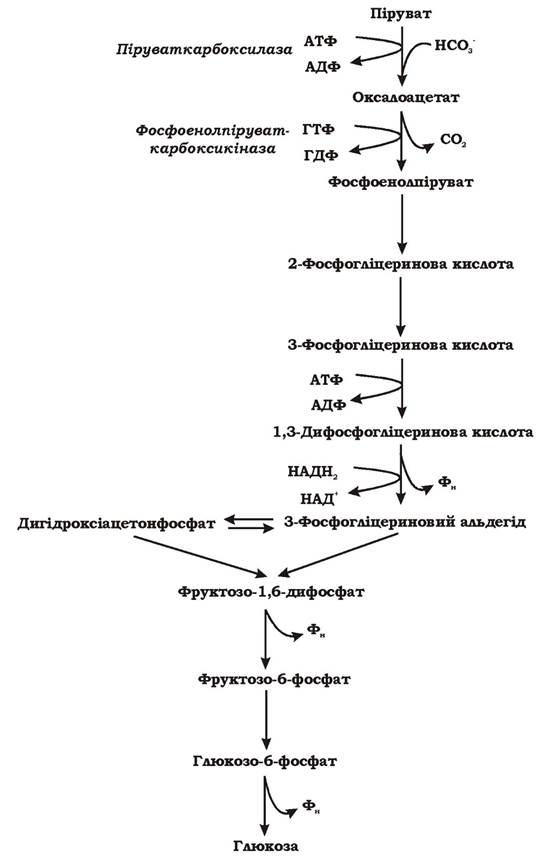
Рис. 5.37. Схема глюконеогенезу
Гліколіз:
Глюкоза + 2 НАД+ + 2 АДФ + 2 Фн → 2 Піруват + 2 НАДН2 + 2 АТФ + 2Н2О
Глюконеогенез:
2 Піруват + 2 НАДН2 + 4 АТФ + 2 ГТФ + 6Н2О → Глюкоза + 4 АДФ + 2 ГДФ + 6 Фн + 2 НАД+ + 2Н+ (ГТФ та АТФ взаємно переходять одна в одну: ГДФ + АТФ → ГТФ + АДФ) Якщо при гліколізі фосфоенолпіруват перетворюється в піруват з виділенням 1 АТФ за одну реакцію під дією піруваткінази, то при глюконеогенезі піруват перетворюється у фосфоенолпіруват за дві реакції, через проміжний продукт ЩОК, з використанням 1 АТФ та 1 ГТФ, під дією двох ферментів: піруваткарбоксилази та фосфоенол-піруват-карбоксикінази.