МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
Розділ 8. МІКРООРГАНІЗМИ І ДОВКІЛЛЯ
ВПЛИВ ФАКТОРІВ ДОВКІЛЛЯ НА МІКРООРГАНІЗМИ
Умови довкілля мають важливе значення для життєдіяльності мікроорганізмів. Серед факторів, які впливають на життєдіяльність мікроорганізмів, виділяють фізичні, фізико-хімічні та хімічні (рис. 8.1).
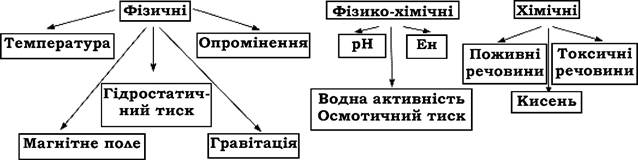
Рис. 8.1. Фактори середовища, що впливають на життєдіяльність мікроорганізмів
Для прокаріотів характерна здатність існувати у значно ширшому діапазоні змін умов середовища, ніж в еукаріотів. Організми широкої
пристосованості називають еврібіонтними (або еврітопними), вузької пристосованості - стенобіонтними (або стенотопними).
Температура. Існування мікроорганізмів значною мірою визначається температурою довкілля. Як і всі живі істоти вони мають свій температурний діапазон, обмежений мінімальними і максимальними значеннями. За впливом температури на мікроорганізми їх поділяють на декілька груп (табл. 8.1): психрофіли, (грец. psychria - холод); мезофіли (грец. mesos - проміжний); термофіли (грец. thermо - тепло).
Таблиця 8.1. Розподіл мікроорганізмів за впливом на них температури
Група мікроорганізмів |
Температура, 0С |
|||
мінімум |
оптимум |
максимум |
||
Психрофіли |
облігатні |
0 і нижче |
5-15 |
20-22 |
факультативні |
0 |
25-30 |
30-35 |
|
Мезофіли |
10-15 |
30-45 |
35-47 |
|
Термофіли |
40-45 |
55-75 |
60-105 |
|
Психрофіли не є рідкісними формами з огляду на те, що полярні регіони займають лише 14 % поверхні Землі. Мікроорганізми, які не лише здатні витримувати низьку температуру, але й надають їй перевагу для свого розвитку, виявляються у ґрунті різних широт, водах морів та океанів, де температура на дні коливається в межах 1-2 оС, а на поверхні - до 5 оС. Психрофільні мікроорганізми поділяють на облігатні і факультативні.
В облігатних психрофілів (Vibrio marinus, Bacillus psychrophilus, Spirillum pleomorphum) мінімальна температура розвитку становить близько 0 оС і нижче, максимальна - 20 оС. Оптимальною температурою для життєдіяльності цієї групи організмів є 5-15 оС, при 25о С вони гинуть. Облігатні психрофіли виділено із сухих долин південної Антарктиди та з арктичних морів.
До факультативних психрофілів відносять мікроорганізми, температурний оптимум яких майже такий самий, як мезофілів, але вони здатні розвиватись і при температурі 0 оС. Психрофіли знайдено серед бактерій Acinetobacter, Aerobacter, Azotobacter, Alcaligenes, Aeromonas, Arthrobacter, Bacillus, Chromobacterium, Clostridium, Cytophaga, Corynebacterium, Flavobacterium, Micrococcus, Nitrobacter, Nitrosomonas, Proteus, Pseudomonas, Vibrio) та грибів родів Stenphylium, Cladosporium, Olvularium та ін.
Причини загибелі мікроорганізмів при низьких температурах пов'язують із "застиганням" мембран, втратою ними текучості та зупинкою трансляції через відсутність гідрофобних взаємодій між рибосомами, блокування ферментів.
Серед механізмів низькотемпературної адаптації психрофітів виділяють:
- збільшення вмісту ненасичених жирних кислот фосфоліпідів у складі цитоплазматичної мембрани (у глибоководних психрофілів у мембранах визначаються незвичні для бактерій поліненасичені жирні кислоти);
- зменшення довжини і водночас збільшення галуження ланцюгів жирних кислот фосфоліпідів у складі ЦПМ;
- стабільність ферментних систем, що забезпечується заміною в білках окремих амінокислот.
Більшість відомих видів прокаріотів належить до мезофілів, які мають температурний оптимум 30-45 оС. Мезофільні бактерії значно поширені у природі. Це типові мешканці ґрунтів та представники мікробіоценозу людини. Класичним представником серед мезофілів є E. coli нижня межа росту 10 оС, верхня - 42 оС, оптимальна температура 37 оС. Патогенні для людини і теплокровних тварин мікроорганізми мають температурний оптимум розвитку близько 37 оС.
Мікроорганізми, які розвиваються при температурі 70 оС і вище, називають термофілами. У межах цієї групи виділяють декілька підгруп: термотолерантні, облігатні, факультативні й екстремальні термофіли.
Мікроорганізми, температурний оптимум яких наближається до такого мезофілів (35-50 оС), але які здатні розмножуватись і за вищих значень температури (70 оС), називають термотолерантними. До них належать деякі представники спороутворювальних бактерій, наприклад Clostridium (ростуть у межах 10-70 оС), Desulfotomaculum (температурний діапазон 20-70 оС, оптимум 30-55 оС), гриби родів Aspergillus, Culvularia, Stenfilium.
Факультативні термофіли мають температурний оптимум значно виший, ніж мезофіли, але здатні розвиватися і при температурі 20 оС. Це представники спороутворювальних бактерій: Bacillus stearothermophilus (topt 50-65 оС); Desulfotomacullum nigrificans (topt 55 оС); Clostridium thermocellum (topt 55-60 оС), а також актиноміцети родів Pseudonocardia, Actinomadura, Microbispora, Micropolyspora, Saccharomonospora, Streptomyces, Thermoactinomyces, Thermomonospora, які мають topt > 50 оС.
Із термальних джерел, температура яких досягає 70-90 оС, виділені облігатні термофіли. Особливістю цих мікроорганізмів є те, що вони не здатні розвиватися при температурі нижче 40-50 оС. Відомими термофілами є: Thermus aquaticus (topt70-75 оС), Bacillus acidocaldarius (topt 60-65 оС), B. caldolyticus (topt 65-80 °С), Thermomicrobium roseum (topt 70-75 оС), Thermoanaerobium brockii (topt 70 оС), Dictyoglomus thermophilum (topt 73-78 оС).
Мікроорганізми, температурний оптимум яких наближається або лежить у межах температури кипіння води, відносять до екстремальних термофілів. Представники екстремальних термофілів об'єднані у групу 35 9-го видання "Berge's Manual of Systematic Bacteriology" і представлені паличкоподібними, нитчастими, кулястими, дископодібними грамнегативними бактеріями, які не утворюють спор і температурний оптимум росту яких становить 75-105 оС. Так, у киплячих сольфатарних водах в Ісландії та Італії виявлено паличкоподібні клітини Pyrobaculum islandicum (topt - 100 оС). З підводних каналів гідротерм на морському дні виділено дископодібні бактерії Pyrodictium occultum (topt - 105 оС). Конічні утворення з сульфідів металів у рифових зонах серединно-океанічних хребтів, так звані чорні курильники, виливають термальні розчини, температура яких досягає 350 оС і які не закипають через високий тиск. Їхні біоценози ще мало вивчені, але в них знайдені паличкоподібні бактерії, які також можуть бути прикладом екстремальних термофілів.
Вивчення механізмів, які забезпечують існування мікроорганізмів при високих температурах, становить значний інтерес. Проведені дослідження показали, що термофілію не можна пояснити лише якою- небудь однією властивістю організму. Запропоновано декілька гіпотез для з'ясування природи термофілії:
- наявність дибіфітаніл-дигліцеринтетраефірів замість фосфоліпідів у мембранах архебактерій;
- збільшення вмісту насичених жирних кислот у фосфоліпідах у складі цитоплазматичної мембрани;
- збільшення довжини і водночас збільшення галуження ланцюгів жирних кислот фосфоліпідів у складі ЦПМ;
- термостабільність ферментів, яка забезпечується заміною в білках окремих амінокислот для підсилення внутрішньомолекулярних взаємодій; стабілізацією ферментів у присутності іонів Mg2+, NH4+,Ca2+;
- здатність синтезувати ферменти подібної субстратної специфічності, але з різним температурним оптимумом;
- синтез білків теплового шоку;
- стабілізація ДНК та РНК у присутності поліамінів (путресцину, спермідину, сперміну), низьких концентрацій Mg2+.
Загалом при високотемпературній адаптації суттєвим є зниження конформаційної гнучкості макромолекул (білків, ліпідів, нуклеїнових кислот), у той час як при низькотемпературній адаптації - навпаки, збільшення їхньої конформаційної гнучкості.
Опромінення. Усі біологічні об'єкти перебувають під впливом різних видів опромінення. Ефекти, які викликаються опроміненням живих організмів, залежать від довжини хвилі світла та його дози, тобто від енергії та кількості квантів, що поглинаються. Електромагнітні хвилі складаються з (рис. 8.2):
- радіохвиль, які мають найбільшу довжину хвилі (понад 1 500 нм);
- світлових хвиль, серед яких виділяють інфрачервоні (до 760 нм), видимі (від 760 до 380 нм), ультрафіолетові (від 380 до 200 нм);
- іонізуючого опромінення (до 200 нм), зокрема рентгенівського (головним чином штучного походження), та γ-хвиль (утворюються при розпаді радіоактивних речовин), космічних хвиль.
Радіохвилі не здійснюють біологічного впливу на мікроорганізми.
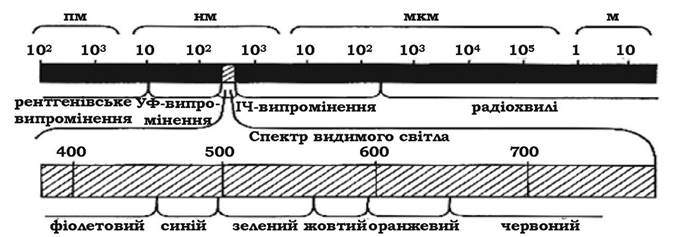
Рис. 8.2. Спектр електромагнітного випромінювання
Інфрачервоні хвилі містять таку незначну кількість енергії, що вони не здатні викликати хімічних перетворень у матерії, яка їх поглинає. Енергія інфрачервоних хвиль перетворюється в тепло.
Випромінювання в ділянці видимого світла забезпечує можливість здійснення впорядкованих реакцій при його поглинанні певними системами. В організмах це опромінення індукує такі процеси, як фотосинтез, фототаксис, фотореактивацію ДНК, синтез деяких макромолекул.
Ультрафіолетове опромінення має сильний бактерицидний ефект. Як дальній ультрафіолет (220-300 нм), так і ближній (300-380 нм) мають пошкоджуючу дію на біополімери клітини в результаті фотолізису, утворення піримідинових димерів, гідроксилювання нуклеотидів, індукції заміни пар основ, однониткових та двониткових розривів ДНК, зшивки ДНК з білками.
Іонізуюче випромінювання є компонентом природної радіації, яка складається переважно з корпускулярного α-, β-опромінення, а також рентгенівського та γ-випромінювання. Радіаційний вплив забезпечують космічні промені, нестабільні ізотопи ґрунту, опадів, розробка копалин радіоактивних мінералів, штучні джерела радіації (випробовування атомної зброї, атомні електростанції, застосування радіоізотопів із медичною та науковою метою).
Дія іонізуючого опромінення на клітини мікроорганізмів виявляється в численних пошкодженнях, подібних до дії ультрафіолету, переважно однониткових та двониткових розривах ДНК. Опосередкований вплив опромінення пов'язаний з утворенням вільних радикалів, що сприяють перекисному окисненню ліпідів.
Мікроорганізми є найбільш стійкими до радіаційного впливу порівняно з іншими організмами. Летальною дозою для ссавців є рівень, наближений до 103 рад, для комах - 105 рад, для мікроорганізмів - 10б рад. Серед мікроорганізмів радіорезистентність коків вища за таку в паличкоподібних, проте серед високорезистентних бактерій відомі як коки (Deinococcus radiodurans, D. radiophilus), так і палички (Deinobacter grandis). Стійкість до опромінення притаманна також різним грибам родів Alternaria, Aureobasidium, Stemphylium, Stachybotrys, Cladosporium.
Місцями існування радіорезистентних мікроорганізмів є радонові джерела, охолоджувальні системи атомних електростанцій, об'єкт укриття атомного реактора.
Механізми радіорезистентності пов'язані із системами репарації, до яких належить ексцизійна репарація (видалення пошкоджених ділянок ДНК) та рекомбінаційна репарація (заповнення дефектів у нуклеотидній послідовності пошкодженої ДНК). Особливістю мікроорганізмів є також контроль систем репарації багатьма генами.
Головним джерелом природного опромінення є сонячна радіація. Основна частина сонячної енергії припадає на долю видимих променів (близько 75 %), майже 20 % - на інфрачервону частину спектра і лише 5 % - на ультрафіолет.
Гравітація. Усі організми зазнають впливу земного тяжіння. Проте доказів прямої дії гравітації на бактерії не існує. Розміри бактеріальних клітин настільки малі, що у водних розчинах вони практично перебувають у стані невагомості, хоча завислі мікроорганізми все ж поступово осідають. Деякі аеробні бактерії у стані невагомості ростуть краще. Це пояснюється більш рівномірним розподілом поживних речовин.
Ще одним прикладом впливу гравітації на мікроорганізми може бути геотропізм, який проявляється у формуванні актиноміцетами і мікроскопічними грибами різнонаправленого повітряного та субстратного міцелію.
Магнітне поле. Магнітне поле помірно впливає на більшість мікроорганізмів неспеціалізованих форм. Доведена стимуляція росту для Pseudomonas aeruginosa, Staphylococcusepidermidis, Halobacterium salinarium магнітним полем із напруженістю 12·103А/м та їх пригнічення магнітним полем з напруженістю 24·103 А/м і вище. Проте у природних середовищах магнітні поля такої напруженості не зустрічаються.
Значний вплив магнітного поля зареєстрований на спеціалізовані форми мікроорганізмів - магнітобактерії. Вони здатні регулювати напрямок свого руху відповідно до напрямку силових ліній магнітного поля. Це явище було названо магнітотаксисом. У слабких магнітних полях (8-40 А/м) магнітобактерії рухаються зі швидкістю до 70 мкм/с уздовж ліній поля.
Магнітобактерії поширені в морських маршах, заболочених прісноводних водоймах, у збірниках для очистки стічних вод, тобто в місцях з малорухомою водою. Чисельність їх у таких умовах досягає 106107 клітин/мл.
Із магнітобактерій найкраще вивченим є Aquaspinllum magnetotacticum - мікроаерофільний хемогетеротроф. Його особливість полягає в наявності магнітосом - кубічних або октаедричних кристалів окису заліза - магнетиту, за рахунок якого вміст заліза у клітинах таких спірил досягає 3,8 %, що набагато перевищує його вміст у клітинах не- спеціалізованих форм (0,025 %). Магнітосоми можуть накопичуватися в осадах водойм у вільному стані, утворюючи біогенні магнетити з високим рівнем природного залишкового магнетизму.
Гідростатичний тиск впливає на мікроорганізми за умови значної амплітуди змін показників цього фактора середовища. Так, в експерименті рівень тиску в 100 атм пригнічував, а в 200 атм стимулював ріст Escherichia coli, однак при 400 атм швидкість росту суттєво зменшувалась, утворювались ниткоподібні клітини.
У природних умовах високий гідростатичний тиск (300-600 атм) створюється на глибині 3 000-6 000 м в океанах. Мікрофлора таких глибин представлена баротолерантними (грец. baros - тягар) мікроорганізмами.
Один відсоток поверхні Землі міститься в океані на глибині понад 10 000 м, де тиск може досягати 1 100 атм. Бактерії, вилучені з таких глибин, зокрема Маріанської западини (11 022 м), не витримують нормального атмосферного тиску і гинуть протягом 5 год. Такі барофільні мікроорганізми вимагають спеціальних культиваторів для підтримання високого тиску.
Механізми баротолерантності і барофільності пов'язують із збереженням текучості мембран, що забезпечується своєрідним жирнокислотним складом ліпідів мембран, збільшенням вмісту довголанцюгових поліненасичених жирних кислот, низькою температурою плавлення полієнових кислот.
Ці механізми подібні до механізмів низькотемпературної адаптації, що сприяє прояву барофілами психрофільних властивостей.
В умовах тиску, який значно нижчий за атмосферний, життєздатність мікроорганізмів зазвичай не порушується.
Вологість. Для забезпечення життєдіяльності всім організмам потрібна вода. Обмін речовин відбувається у водних розчинах. Вода, яка міститься в зовнішньому для мікроорганізму середовищі, може бути доступною або недоступною для нього. Доступність води для мікроорганізмів найчастіше виражають через водну активність, яка корелює з тиском водяної пари над розчином і обчислюється при вимірюванні відносної вологості повітря (Ww):
![]()
де Р - тиск водяної пари над розчином, Ро - тиск пари над водою. Наприклад, над 35 % розчином сірчаної кислоти при температурі 25 оС тиск пари в міліметрах ртутного стовпчика становить 15,8, а тиск пари над водою - 23,76. Тоді Ww дорівнює 67 %о. Активність води - це та ж відносна вологість, але не виражена у відсотках, тобто якщо Ww = 67 %, то Aw = 0,67.
Різні види мікроорганізмів вимагають певного рівня водної активності. При мінімальній водній активності (0,8-0,6) можуть існувати мікроскопічні гриби (Penicillium, Aspergillus, Cladosporium та ін.). Максимальної водної активності (1,0-0,9) потребують нейстонні бактерії
родів Spirillum, Nevskia, Hyphomicrobium, Caulobacter, сульфат- редуктори Desulfotomaculum та ін.
Активність води може бути також виражена через концентрацію розчинених речовин:
![]()
де П1 - кількість молей розчинника, П2 - кількість молей розчиненої речовини.
З рівняння видно, що підвищення концентрації розчинених речовин знижує водну активність, а отже і доступність води для клітини.
Концентрація речовин, розчинених у зовнішньому для мікроорганізму середовищі, зумовлює осмотичний тиск середовища, який перебуває в оберненій залежності від водної активності.
Надходження води у клітину можливе, якщо осмотичний тиск у клітині більший, ніж тиск зовнішнього розчину. Нормальний тиск у клітині зазвичай лежить у межах 3-6 атм, що відповідає такому середовищу існування як, наприклад, ґрунт (0,5-5 атм). У солоних середовищах або з високим вмістом цукрів (варення, мед) осмотичний тиск може досягати 100 атм. Мікроорганізми уникають осмотичного стресу завдяки наявності системи осморегуляції, складовими якої є:
- білки осмосенсорів, які реагують на зміну осмотичного тиску;
- зміна співвідношення білків поринів, що утворюють гідрофільні пори;
- синтез осмопротекторів (високорозчинних речовин, концентрація яких врівноважує зовнішній тиск, - аміномасляна, глютамінова кислота, пролін);
- система вибіркового накопичення іонів у клітині - збільшення концентрації К+ при збільшенні зовнішнього тиску);
- система регуляції осмолярності периплазми - синтез олігосахаридів, що утримують катіони).
Не тільки осмотичний тиск зменшує доступність води для клітини. Існує інший шлях - дефіцит екзогенної води, що може призводити до матричного водного стресу, до якого мікроорганізми більш чутливі, ніж до осмотичного.
Пристосуванням мікроорганізмів в умовах дефіциту екзогенної води є спороутворення, утворення гігроскопічних слизових капсул, здатність до використання метаболічної води в результаті окиснення органічних сполук (з 100 г глюкози утворюється 60 г води).
Реакція середовища (рН). Для життєдіяльності мікроорганізмів велике значення має реакція середовища, яка визначається концентрацією іонів водню, що утворюються у водному розчині при електролітичній дисоціації води. Кількісно реакцію середовища виражають за допомогою рН-показника. Цей показник означає величину від'ємного логарифму концентрації іонів водню. Концентрація іонів водню чистої води дорівнює 10-7, тобто рН = 7. Рівні рН нижче 7 визначають кислотність середовища, понад 7 - її лужність.
Кожний мікроорганізм існує в певних межах рН і має свій специфічний оптимум реакції середовища (рис. 8.3).
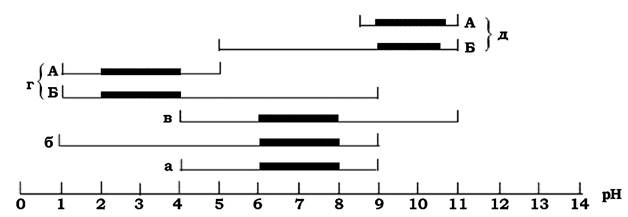
Рис. 8.3. Межі та оптимальні значення росту прокаріотів залежно від pH:
а - нейтрофіли; б - кислотостійкі; в - лугостійкі; г - ацидофіли; д - алкалофіли. А - облігатні; Б - факультативні форми.
Жирна лінія - оптимальні pH росту
Ацидофіли (лат. addus - кислий) існують у межах рН від 1,0 до 6,0 і представлені термофільними архебактеріями та еубактеріями, наприклад: Thermoplasma addophilum (pH 1,8), Sulfolobus acidocaldarius (pH 2,0-3,0), Badllus addocaldarius (pH 3,0), Sulfobacillus thermosulfidooxidans (pH 2,0) і мезофільними хемолітоавтотрофами, наприклад: Thiobadllusacidophilus (pH 3,0-3,5), T. ferrooxidans (pH 1,8-2,4), T. thiooxidans (pH 2,5), T. thioparus (pH 3,0).
Алкалофіли (араб. алькалі - луг) поширені в середовищах з рН 8,5-11,5 (у ґрунтах, місцях розкладу білків, біологічних рідинах із лужною реакцією). Так, Bacillus pasteuni потребує для свого розвитку pH 9,0, Sporosarcina urea,e - pH 9,0, B. firmus - pH 9,0-10,5, B. alcalophilus - pH 10,5.
Вплив іонів водню на мікроорганізми може бути як прямим, так і опосередкованим. Опосередкований вплив пов'язаний із впливом іонів водню на певні компоненти середовища, дисоціація яких залежить від рН і відбивається на надходженні поживних речовин у клітину. Реакція середовища здійснює вплив на утворення та активність мікробних ферментів.
Окисно-відновний потенціал (Eh). Природні середовища існування мікроорганізмів характеризуються також окисно-відновним потенціалом (Eh), який відображає співвідношення окиснених та відновлених сполук у біологічних системах і є індикатором біохімічних процесів.
![]()
де Е0 - потенціал електрода порівняння; R - газова константа (8,3144 Джтрад-1-моль-1; за 25 оRT = 2,479 кДж-моль-1); F - число Фарадея (1F = 96 485 Кл-моль-1); Т - абсолютна температура; Cred - концентрація відновлених сполук; Сoх - концентрація окиснених сполук.
Вимірюється Eh у мілівольтах і має найвище позитивне значення за високих концентрацій окисника (+815 мВ - реакція розкладу Н2О на О2 і Н2) та найбільше негативне значення при високих концентраціях відновників (-414 мВ - реакція відновлення атомарного водню до молекулярного).
Високоаеровані середовища з рівнем Eh +810 мВ створюють умови для існування облігатних аеробів та деяких факультативно анаеробних мікроорганізмів родів Pseudomonas, Bacillus, Corynebacterium, Arthrobacter, Streptomyces. Факультативні анаероби переважають у середовищах, окисно-відновний потенціал яких наближається до нуля.
Для нітратредукуючих та денітрифікуючих бактерій видів Pseudomonas fluorescens, Bacillus licheniformis, Paracoccus denitrificans, Thiobacillus denitnficans характерний Eh +440 мВ і нижче. Ці мікроорганізми поширені у ґрунті, де здійснюють важливі перетворення в циклі азоту. Нітратредукуючі бактерії Escherichia, Enterobacter, Proteus тощо є представниками мікрофлори шлунково-кишкового тракту, нижні відділи якого переважно заселені анаеробними бактеріями родів Bacteroides, Propionibacterium та ін.
Облігатні анаероби існують у середовищах з високими негативними значеннями Eh. Серед них відомі метаногенні бактерії (Eh - 220 мВ) з природних (лимани, болота, рисові поля, рубець жуйних тварин) та антропогенних еконіш (відстійники очисних споруд) - Methanobacterium thermoautotrophicum, Methanosarcina barkeri, Methanococcus mazei, Methanospirillum hungatii.
Сульфатредукуюча бактерія Desulfotomaculum ruminis також є представником мікрофлори рубця жуйних тварин, проте більшість сульфатредукторів (Eh -265 мВ) становлять біоценози мулу озер та морських осадів, де створюються анаеробні умови, наприклад: Desulfovibrio desufuricans, Desulfosarcina variabilis, Desulfococcus multivorans, Desulfonema limicola, Desulfobacter curvatus, Desulfotomaculum nigrificans. Ці мікроорганізми також мають значення як корозійнонебезпечні.
Поживні речовини. Поживними речовинами є будь-які сполуки, використані мікроорганізмами як джерела макро- та мікроелементів, електронів та енергії. Надходження поживних речовин відбувається нерівномірно в часі та просторі, тому прийнято характеризувати місця існування мікроорганізмів термінами усередненого потоку поживних речовин за добу.
Бактерії, існування яких у природі залежить від їхньої здатності розмножуватись у місцях з низьким потоком поживних речовин - до 0,1 мг/л за добу, відносять до оліготрофів (грец. oligos - малий, trophe - їжа). Типовими представниками оліготрофів є мешканці водойм та ґрунтів (Achromatium beggiatoa, Thiothrix beggiatoa, Thiothrix та ін.).
Копіотрофами (грец. copiosus - надмірність, trophe - їжа) називають мікроорганізми, що надають перевагу багатим субстратам. Копітрофи розвиваються в умовах надходження поживних речовин, які щонайменше в 50 разів перевищують такі, що сприяють розвитку оліготрофів. До копітрофів належать E. coli, бактерії роду Zymomonas та інші мікроорганізми, виділені з середовищ з активними процесами гниття.
Оліготрофні мікроорганізми досить поширені у природі і в багатьох випадках чисельно домінують над копітрофами. Поширенню оліготрофів сприяє ряд їхніх особливостей, а саме:
- накопичення резервних речовин (полі-β-оксимасляної кислоти, по ліглюкози, поліфосфатів);
- транспортні системи функціонують в умовах мінімальних концентрацій поживних речовин і характеризуються низькою субстратною специфічністю, нездатністю дискримінувати речовини;
- здатність до мініатюризації клітин в умовах голодування;
- низька швидкість розмноження - час генерації оліготрофів становить 20-200 год на противагу копіотрофам, які утворюють нове покоління протягом 10-20 хв.
Досить часто мікроорганізми, виділені як оліготрофи, під час лабораторного культивування стають копітрофами.
Шляхи адаптації до джерел живлення можуть бути спеціалізованими або неспеціалізованими. Мікроорганізми, які здатні використовувати вузький спектр субстратів або вимагають для росту конкретну, незамінну іншими, речовину, відомі як спеціалісти. До них належать, наприклад, тіонові бактерії. Такі мікроорганізми швидше ростуть в умовах надлишку доступного для них субстрату. Генералісти використовують різноманітні сполуки, тому легше знаходять джерела живлення. Серед оліготрофів переважають генералісти, хоча є і спеціалісти, як, наприклад, метилотроф Hyphomicrobium vulgare.
Токсичні речовини. Можливість нормального існування тих чи інших бактерій залежить не лише від доступності поживних речовин, а й від їх можливості протистояти згубній дії токсичних для них речовин. Рідкісне явище у природних еконішах - це повна відсутність токсичних речовин.
За типом впливу токсичних речовин на клітину їх дія може бути бактерицидною або бактеріостатичною. Бактеріостаз (грец. bacterion - паличка, stasis - стояти на місці) - затримка росту і розмноження бактерій. Припинення дії фактора приводить до відновлення росту. При бактерицидності (лат. caedere - вбивати) дія фактора призводить до загибелі клітини.
У багатьох випадках речовина в малих концентраціях буде мати бактеріостатичний ефект, тоді як у високій - бактерицидний.
За характером впливу хімічні речовини поділяють на такі, що призводять до:
- пошкоджень клітинної стінки (поверхнево-активні речовини, жирні кислоти, мила, детергенти);
- пошкоджень клітинної стінки та білків цитоплазми (фенол, крезол та їхні похідні);
- порушень процесів клітинного ділення завдяки спорідненості до нуклеїнових кислот (акридини);
- денатурації білків (формальдегіди);
- коагуляції білків (солі важких металів).
Присутність токсичних речовин зменшує можливості збалансованого обміну між клітинами мікроорганізмів та середовищем. Це активізує контролюючі та захисні системи мікроорганізмів, до яких належать:
- система строгого контролю, що регулюється синтетазою фосфонуклеотидів-алармонів (франц. alarme - тривога) і спрямована на обмеження поглинання сполуки із середовища;
- система SOS-відповіді, яка активує каскадні реакції дерепресії білків з протеолітичною активністю, репараційних систем, зміни складу білків зовнішньої мембрани клітин бактерій;
- система адаптивної відповіді - індуцибельна антимутагенна система репарації, що активується концентраціями мутагенів у 10100 разів менших, ніж необхідно для прояву їхньої мутагенної дії;
- синтез білків теплового шоку, який відбувається не тільки в умовах температурного стресу, але й під впливом опромінення та токсичних речовин;
- синтез ферментів захисту від окисників - каталази, пероксидази, супероксиддисмутази, глутатіонредуктази, алкілгідропероксидре- дуктази, глюкозо-6-фосфатдегідрогенази.
- формування резистентності шляхом активної депортації токсичних
сполук із клітини, синтезу гідролаз, редуктаз, трансфераз.
Мікроорганізми здатні концентрувати токсичні речовини в поверхневих структурах і навіть всередині клітини. Коефіцієнти концентрування різні для різних мікроорганізмів та сполук. Іноді накопичення таких речовин є летальним для клітини, в інших випадках - ні. Цю властивість мікроорганізмів використовують з практичною метою в технологіях біосорбції, трансформації сполук важких металів, детоксикації природних середовищ.
Важливою адаптаційною ознакою є здатність певних мікроорганізмів використовувати речовини, відомі своєю токсичною дією, як джерела енергії. Так, серед нітрифікуючих бактерій відомі Nitrobacter, Nitrospina, Nitrococcus, Nitrospira, єдиним джерелом енергії яких слугує нітрит у реакції його окиснення до нітрату. Така властивість цих мікроорганізмів забезпечується нітритоксиредуктазною активністю. Бактерії роду Nitrosomonas, Nitrosococcus, Nitrosospira, Nitrosolobus окиснюють аміак до нітриту завдяки активності ферментів монооксигена- зи та гідроксиламіноксидоредуктази. Сіркоокисню-вальні бактерії родів Thiobacillus, Thiomicrospira, Thiosphaera, Thermothrix використовують як джерело енергії відновлені сполуки сірки - потенційно токсичні субстрати: сірководень, тіоціанат, сульфіти.
Деякі мікроорганізми, наприклад коринебактерії, псевдомонади, здатні утилізувати продукти нафтопереробки, нафталін, фенол тощо.
Кисень. Сучасний стан атмосфери Землі, яка містить ~21 % кисню, зумовив формування певних груп організмів за впливом на них цього газу. Деякі мікроорганізми обрали місця існування, де О2 практично відсутній, і тим самим забезпечили собі умови "безкисневої епохи» - це облігатні анаероби (Desulfotomaculum, Syntrophospora). Інші - облігатно аеробні мікроорганізми пристосувались до кисневих умов і активно використовують кисень у своїх метаболічних процесах, наприклад Acetobacter. Факультативні анаероби мають альтернативні шляхи енергетичного обміну і можуть розвиватися як за наявності, так і у відсутності кисню, зокрема Eschenchia, Proteus. Деякі аеробні мікроорганізми потребують для свого розвитку молекулярний кисень, але у значно меншій концентрації, ніж він міститься в атмосфері. Їх називають мікроаерофілами, наприклад Campylobacter, Helicobacter. Виділяють також аеротолерантні мікроорганізми, які мають бродильний тип метаболізму, але при контакті з киснем не гинуть.
Дія молекулярного кисню на клітину досить агресивна і пов'язана з його здатністю окиснювати клітинні метаболіти, що не є бажаним для
клітини. Крім того, при окисненні флавопротеїдів утворюється пероксид водню - токсична для бактерій речовина. У реакціях окиснення утворюється ще більш токсичний вільний перекисний радикал - O2-, хоча і в невеликій кількості.
Сучасні аеробні та аеротолерантні прокаріоти для боротьби з токсичними формами кисню виробили захисні механізми, основою яких є здатність спеціальних ферментів каталізувати реакції їхнього руйнування. Наприклад, супероксиддисмутаза запобігає накопиченню потенційно летального для прокаріотів перекисного радикалу, каталізуючи його перетворення на кисень та перекис водню. Каталаза, у свою чергу, каталізує розпад перекису водню на кисень та воду:
Ці ферменти містяться лише у клітинах аеробів. В аеротолерантних та факультативних анаеробів, як правило, каталаза відсутня. У клітинах анаеробних мікроорганізмів не виявлено жодного з цих ферментів, тому вони беззахисні проти токсичної дії кисню. Єдино досяжні для них шляхи нейтралізації кисню - витіснення його з середовища існування завдяки інтенсивному виділенню СО2 та Н2.
У природних умовах неможливо відокремити вплив на мікроорганізми якогось одного фактора. Сукупність впливу факторів може різко впливати на дію досліджуваного. Так, рН середовища змінює летальний вплив температури. Мікроорганізми швидше гинуть при нагріванні в кислому середовищі, ніж нейтральному або лужному. Летальний вплив рентгенівських променів значно підвищується у присутності молекулярного кисню. Тобто терміни "оптимальна температура» або "оптимальний рН" мають реальне значення, коли відомі всі інші фактори довкілля.