МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
Розділ 9. РОЛЬ МІКРООРГАНІЗМІВ У КРУГООБІГУ РЕЧОВИН У ПРИРОДІ
Кругообіг заліза
Залізо в невеликих кількостях входить до складу клітин. Цей елемент визначається у структурі деяких ферментів (каталаза, нітрогеназа, пероксидаза), цитохромів, залізопорфіринових сполук та ін.
У ґрунті залізо міститься у формі органічних і неорганічних сполук. Рослини здатні асимілювати лише мінеральні, розчинні у ґрунтовій воді, форми заліза. Роль ґрунтових мікроорганізмів полягає в тому, що вони трансформують нерозчинні (недоступні для рослин) сполуки заліза у водорозчинні і навпаки. Біологічний цикл заліза наведено на рис. 9.7.
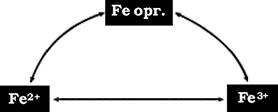
Рис. 9.7. Біологічний цикл заліза
При мінералізації органічних сполук заліза мікроорганізми діють на органічну частину молекули, асимілюючи її, залізо при цьому звільняється і в аеробних умовах осаджується у вигляді гідрату його оксиду, тобто не використовується мікробною клітиною для метаболічних процесів. Таку здатність проявляють хемоорганотрофні мікроорганізми, серед яких є бактерії і гриби.
Деякі мікроорганізми здатні окиснювати відновлені сполуки заліза. Гідрат оксиду заліза, який при цьому утворюється, відкладається на поверхні мікробної клітини, утворюючи часто чохли, інкрустовані залізом. Це хемоорганотрофи забруднених водойм (бактерії роду Blastocaulis) і ґрунтів (представники родів Hyphomicrobium, SelibeHa та ін.).
Бактерії роду Hyphomicrobium представлені клітинами овальної або бобоподібної форми (0,3-1,2 х 1,0-3,0 мкм), які мають полярні про- стеки різної довжини й діаметром 0,2-0,3 мкм. Розмноження відбувається брунькуванням, брунька при цьому утворюється на вершині простеки. Зріла брунька, яка відділяється від материнської клітини, має один або декілька полярних чи субполярних джгутиків (рис. 9.8).
Це аероби і хемоорганотрофи (H. coagulans, H. vulgare, H. zavarzinii та ін.).

Рис. 9.8. Електронна мікрофотографія Hyphomicrobium sp.: зріла клітина, видно гіфи і термінальні бруньки
Рід SelibeHa представлений клітинами паличкоподібної форми (0,5-0,8 х 1,0-12,0 мкм) з одним субполярним джгутиком, покритим чохлом або декількома латеральними джгутиками без чохлів (рис. 9.9). Полярний ріст подовжених клітин представників цього роду в результаті асиметричного поділу супроводжується утворенням коротких клітин-швермерів і довгих нерухомих клітин. Це облігатні аероби, гетеротрофи (S. stellata).
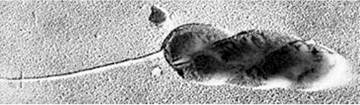
Рис. 9.9. Електронна мікрофотографія Seliberia sp.: видно спірально закручену клітину і джгутик
Мікроорганізми здатні також окиснювати неорганічні сполуки заліза в болотах, дренажних трубах, озерах та інших водоймах з утворенням вохристих осадів. Цей процес здійснюється групою залізобактерій, до яких належать представники родів Gallionella, Metallogenium, нитчасті бактерії родів Leptothrix, Crenothrix та ін.
Представники роду Gallionella мають паличкоподібні клітини (0,50,7 х 0,8-1,8 мкм). Виділяють колоїдний гідроксид заліза (феригідрит) з утворенням стеблинок, форма й структура яких служить основною діагностичною ознакою. Клітини розташовуються апікально на кінці стеблинки, перпендикулярно до її осі. Рухаються за допомогою єдиного полярно локалізованого джгутика. Аероби, мікроаерофіли, хемолітоавтотрофи. Донором електронів служить виключно Fe2+. Урожай клітин становить 1 г сухої біомаси на 150 г окисненого двовалентного заліза. Це одні з найважливіших залізобактерій, які утворюють значні кількості феригідриту у водоймах та системах водопостачання (G. furriginea).
Рід Metallogenium представлений кулястими клітинами (0,2-1,5 мкм), які позбавлені клітинної стінки. Це мікоплазмоподібні організми, що проростають безпосередньо або шляхом повторного брунькування створюють групу округлих "елементарних тіл", при проростанні яких утворюється одна або декілька гнучких ниток. Стадія життєвого циклу має коки і коки з нитками, які звужуються і розходяться радіально від центра і покриті оксидами. Нитки (або араї) мають товщину близько 0,02-0,3 мкм. Мікроколонії (або ценобії) мають один або декілька центральних коків і тонкі араї, які відходять від центра у формі "павучків", що дає можливість легко їх розпізнавати на цій стадії. Це хемоорганотрофи або паразити грибного міцелію, аероби (M. personatum).
Бактерії роду Leptothrix утворюють нитки з чохлами, інкрустованими оксидами. Нитки нерухомі, але можуть утворюватися рухомі поодинокі клітини, які мають джгутики. Клітини паличкоподібні (0,6-1,4 х 1,0-12,0 мкм), утворюють ланцюжки, покриті чохлом, або перебувають у вільному стані. Чохли просочені або покриті оксидами заліза й мангану (рис. 9.10). Часто виявляються порожні чохли без клітин. За типом живлення це хемоорганотрофи з дихальним типом метаболізму. Діапазон температур від 10 оС до 35 оС, а оптимум - у межах 25 оС. Оптимальне значення pH 6,5-7,5. Типовий вид - L. ochraceae. Бактерії цього роду окиснюють двовалентне залізо (FeCO3) до тривалентного Fe2(CO3)3з наступною його трансформацією до Fe(OH)3:
![]()

Рис. 9.10. Leptothrix pseudo-ochracea:
Представники роду Crenothrix мають клітини від циліндричної до дископодібної форми. Поділ клітини відбувається шляхом утворення поперечних перегородок з утворенням ниток до 1 см завдовжки, які покриваються чохлами. Нитки часто прикріплюються до твердого субстрату (рис. 9.11). Чохол, який оточує нитку, досить тонкий, на вершині може бути безбарвним або, в основі, інкрустується оксидом заліза (або мангану). На кінцях ниток спостерігаються потовщення, в яких утворюються сферичні клітини розмноження - "макрогонідії", які висковзують із чохла й утворюють нові нитки з чохлами. У потовщені нитки поділ клітини може відбуватися в поздовжній і поперечній площинах.

Рис. 9.11. Crenothrix polyspora
Дрібні кубічні клітини ("мікрогонідії"), які утворюються при цьому, заокруглюються, висковзують і утворюють нові нитки. Деякі ланцюжки клітин проявляють ковзний тип руху. Бактерії роду Crenothrix виявляються у стоячих і проточних водах, які містять незначні концентрації Fe 2+ і метану. Типовий вид - C. polyspora.
Описані також облігатні хемолітоавтотрофні бактерії, які здатні отримувати енергію за рахунок окиснення закисного заліза і використовувати вуглець CO2. До таких бактерій належить сіркоокиснювач Thiobacillius ferrooxidans, який забезпечує перебіг реакцій за такою схемою:
![]()
Окиснення закисних форм заліза дає низький вихід енергії, і для забезпечення свого росту бактерії трансформують значні кількості заліза. Так, для утворення 1 г біомаси бактерії повинні окиснити близько 500 г сірчанокислого заліза.
Установлено також здатність до накопичення окислів заліза деякими фототрофними бактеріями (наприклад, ціанобактеріями), нитчастими зеленими бактеріями і деякими водоростями.
Вважається, що хемолітотрофні і ряд хемоорганогетеротрофних мікроорганізмів, під впливом яких відбувається трансформація заліза у природі, беруть участь в утворенні відкладень заліза, які забезпечують формування родовищ заліза в болотах, озерах та інших водоймах.