ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 1. Общие сведения об иммунной системе
РАСТВОРИМЫЕ МЕДИАТОРЫ ИММУНИТЕТА
В развитии иммунного ответа участвует целый ряд молекул-посредников, в том числе выделяемые лимфоцитами антитела и цитокины, а также различные белки сыворотки, обычно содержащиеся в ней в низкой концентрации. Эти белки названы острофазными, так как их концентрация быстро нарастает при инфекционном процессе. Один из примеров — это С-реактивный белок (CRP), названный так за способность связываться с С-белком пневмококков. Благодаря такому связыванию фагоциты начинают более активно поглощать пневмококки — процесс, называемый опсониза- цией (см. рис. 1.10). В качестве опсонинов, т. е. опсонизирующих молекул, действуют главным образом антитела и компоненты комплемента.
Белки системы комплемента служат медиаторами фагоцитоза, регулируют воспалительную реакцию и, взаимодействуя с антителами, участвуют в иммунной защите организма
В систему комплемента входят около двух десятков сывороточных белков, общая функция которых состоит в регуляции воспаления. Компоненты комплемента взаимодействуют между собой и с другими элементами иммунной системы. Например, ряд микроорганизмов спонтанно активирует систему комплемента по так называемому альтернативному пути, представляющему собой механизм врожденного, неспецифического иммунитета. В результате с поверхностью микробов связываются компоненты комплемента, что приводит к поглощению этих возбудителей фагоцитами. Когда система комплемента принимает участие в реакциях специфического приобретенного иммунитета, ее обычно активируют по классическому пути антитела, связавшиеся с поверхностью клеток микроорганизма.
Активация комплемента — это каскад реакций, в котором каждый предшествующий компонент действует на последующий, подобно тому, как это происходит при свертывании крови. Как классический, так и альтернативный путь активации комплемента приводит к образованию белков или пептидных фрагментов, вызывающих следующие эффекты:
✵ опсонизация микроорганизмов с последующим поглощением и внутриклеточным разрушением их фагоцитами;
✵ привлечение фагоцитов к месту воспаления (хемотаксис);
✵ усиление тока крови в месте активации и повышение проницаемости капилляровдля компонентов плазмы;
✵ повреждение собственных клеток организмов, грамотрицательных бактерий и вирусов или других микробов, вызвавших активацию комплемента; лизис возбудителя вследствие такого повреждения обеспечивает прекращение инфекции;
✵ дополнительное выделение медиаторов воспаления из тучных клеток.
Эти функции системы комплемента схематически представлены на рис. 1.7 и подробно описаны в гл. 4.
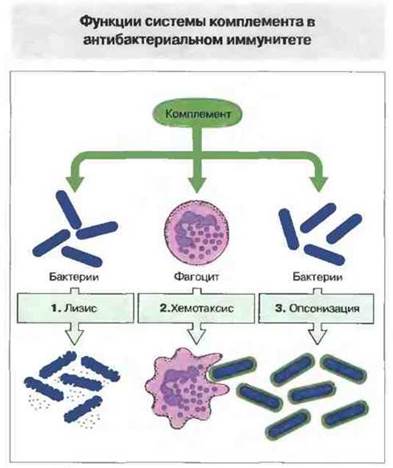
Рис. 1.7. 1. Система комплемента способна без участия антител разрушать мембраны многих видов бактерий. 2. Продукты активации комплемента привлекают к месту реакции фагоциты (хемотаксис). 3. Покрывая в результате связывания поверхность бактерий (опсонизация), компоненты комплемента облегчают фагоцитам их распознавание и поглощение. Все зти реакции могут протекать за счет собственной способности комплемента к распознаванию микробных структур или с помощью антител, связавшихся с бактериями.
Цитокины - разнообразные белки, осуществляющие передачу сигналов между лимфоцитами, фагоцитами и другими клетками организма
Цитокинами обобщенно называют большую группу соединений, участвующих в межклеточной передаче сигналов в ходе иммунного ответа. Все цитокины — это белки или пептиды; некоторые из них представляют собой гликопротеины. Цитокины подразделяют на несколько групп Так, цитокины, выделяемые лимфоцитами, часто называют лимфокинами. Основные группы цитокинов приведены ниже.
Интерфероны (ИФ). Эти цитокины особенно важны в сдерживании некоторых вирусных инфекций. Одну группу интерферонов (ИФα и ИФβ) синтезируют клетки, инфицированные вирусами. Другой тип интерферона (ИФγ) выделяют некоторые активированные Т-клетки. Интерфероны сообщают противовирусную устойчивость незаряженным тканевым клеткам (рис. 1.8). Они образуются на ранней стадии инфекции и создают первую линию защиты против большинства вирусов.
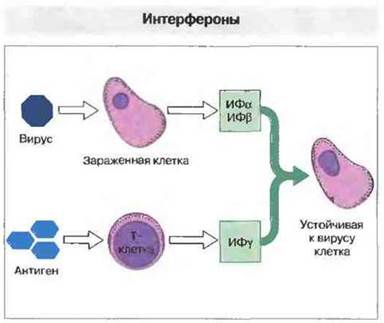
Рис. 1.8. Инфицированные вирусом клетки способны синтезировать интерферон, одни - интерферон-α (ИФα), другие - интерферон-β (ИФβ). Некоторые субпопуляции Т-лимфоцитов после активации антигеном образуют интерферон-γ (ИФγ): Воздействуя на другие клетки организма, иктерфероны сообщают им устойчивость к вирусной инфекции. Интерферон-γ оказывает также много других эффектов.
Интерлейкины (ИЛ). Это большая группа цитокинов (от ИЛ-1 до ИЛ-17), синтезируемых в основном Т-клетками, но в некоторых случаях также мононуклеарными фагоцитами или другими тканевыми клетками. Интерлейкины обладают разнообразными функциями, но большинство их стимулирует другие клетки для деления или дифференцировки. Каждый интерлейкин действует на отдельную, ограниченную группу клеток, экспрессирующих специфичные для данного ИЛ рецепторы.
Колониестимулирующие факторы (КСФ). Эти цитокины участвуют в регуляции деления и дифференцировки стволовых клеток костного мозга и клеток — предшественников лейкоцитов крови. Балансом рахтичных КСФ в определенной мере обусловлено соотношение между различными типами образующихся в костном мозге лейкоцитов. Некоторые КСФ стимулируют дальнейшую дифференцировку клеток и вне костного мозга.
Другие цитокины. Среди них значительную роль играют факторы некроза опухолей (ФНОα и ФНОβ) и трансформирующий фактор роста β (ТФРβ). Они выполняют разнообразные функции, но особенно важны как медиаторы воспаления и цитотоксических реакций.
Антитела специфически связываются с антигеном и затем опосредуют вторичные эффекты
Антитела (Ат), называемые также иммуноглобулинами (lg), — это семейство сывороточных белков, образуемых В-лимфоцитами. Как уже было отмечено выше, они представляют собой растворимую форму антигенсвязываюших В-клеточных рецепторов. Все поликлональные антитела данного изотипа однотипны по строению молекулы, но различаются областью антнгенсвязывающего центра. Как правило, каждое антитело может специфически связываться только с одним антигеном.
В то время как одна часть молекулы антитела (Fab-фрагмент) предназначена для связывания с антигеном, другая его часть (Fc-фрагмент) взаимодействует с различными элементами иммунной системы, например с фагоцитами или с одним из компонентов комплемента. Фактически, антитела действуют как молекулы-посредники, вовлекающие различные элементы иммунной системы в распознавание патогенных микробов и продуктов их метаболизма (рис. 1.9)
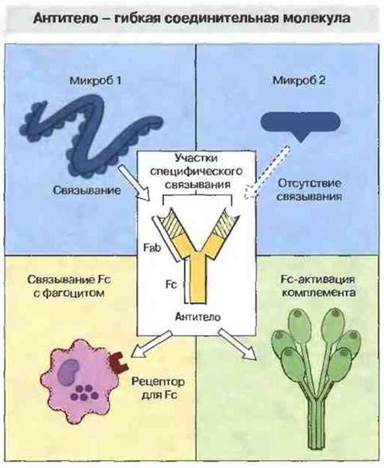
Рис. 1.9. Если микроб не обладает свойством активировать комплемент или связываться с фагоцитами, организм-хозяин образует против него антитела, которые как соединительные молекулы связывают микробные клетки с комплементом и фагоцитами. В организме человека могут синтезироваться несколько миллионов различных по специфичности антител, способных распознавать множество самых разнообразных инфекционных агентов. Например, изображенное на схеме антитело связывается своим антигенсвязывающим фрагментом (Fab) с микробом 1, но не с микробом 2. Fc-фрагмент антитела может активировать комплемент или связываться с рецепторами для Fc на клетках организма-хозяина, прежде всего на фагоцитах.
Та часть молекулы антитела, которая взаимодействует с клетками иммунной системы, названа Fc-фрагмеитом. Нейтрофилы, макрофаги и другие мононуклеарные фагоциты несут на своей поверхности рецепторы для Fc-фрагмента. Следовательно, если антитела связались с патогенным микроорганизмом, они могут своим Fc-фрагментом взаимодействовать и с фагоцитами. Благодаря этому клетки возбудителя будут поглощены (фагоцитированы) и разрушены фагоцитами. Антитела в данном случае действуют как опсонины. Кроме того, фагоциты распознают чужеродный материал, используя в качестве опсонина активированный компонент комплемента (СЗЬ), но наиболее всего фагоцитоз эффективен в присутствии одновременно антител и комплемента (рис. 1. 10).
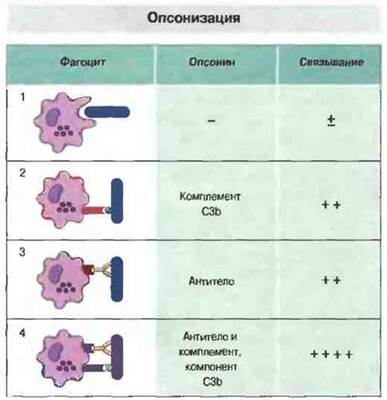
Рис. 1.10. 1. Фагоциты обладают способностью непосредственно, т. е. без участия опсонинов, связывать бактерии и другие микроорганизмы, но при активации микробами комплемента связывание намного усиливается. 2. В этом случае бактерии взаимодействуют с С3b, благодаря чему фагоциты получают возможность связывать их посредством рецепторов для С3b. 3. Для фагоцитоза микробов, которые в полной мере или совсем не активируют комплемент, необходима опсони- зация антителами (Ат), способными связываться с Fc-рецепторами на поверхности фагоцитов. 4. Антитела могут сами активировать комплемент, и если микробы опсонизированы не только антителами, но также С3b, их связывание фагоцитами существенно возрастает.