ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 12. Развитие иммунной системы в онтогенезе
ЛИМФОИДНЫЕ КЛЕТКИ
Недавно проведенные эксперименты на мышах показали, что общий предшественник лимфоидных клеток впервые появляется в каудальной части спланхноплевры. Клетки-предшественники, вероятно, мигрируют с кровотоком в желточный мешок, а затем в первичные лимфоидные органы — тимус и печень плода, где они развиваются соответственно в Т- и В-клетки. Зрелые лимфоциты перемещаются затем во вторичные лимфоидные ткани, где приобретают способность реагировать на антиген.
Т-клетки развиваются в тимусе
Образование Т-клеток начинается с миграции стволовых клеток. Тимус развивается из третьего (а у некоторых видов и из четвертого) глоточного кармана в виде эпителиального зачатка эндо- и эктодермального происхождения, который заселяется стволовыми клетками из крови. Для формирования огромного разнообразия зрелых Т-клеток с различной специфичностью антигенных рецепторов требуется, по-видимому, относительно немного стволовых клеток. В образовании закладки тимуса, по крайней мере у мыши, участвуют два слоя эмбриональной ткани: эктодерма третьей жаберной щели, из которой формируется эпителий корковой зоны тимуса, и эндодерма третьего глоточного кармана, дифференцирующаяся в эпителий мозговой зоны тимуса.
Как показывают экспериментальные исследования, миграция стволовых клеток в тимус происходит не случайно, а в ответ на хемотаксические сигналы, периодически исходящие из зачатка тимуса. Одним из хемоаттрактантов может служить β2-микроглобулин, компонент молекул МНС класса I. У птиц колонизация тимуса стволовыми клетками происходит двумя или тремя волнами, но у млекопитающих такой волнообразный процесс не доказан. Попав в тимус, стволовые клетки под влиянием эпителиального микроокружения начинают дифференцироваться в тимические лимфоциты (называемые тимоцитами). Неясно, являются ли стволовые клетки «пре-Т-клетками», т. е. начинается ли их дифференцировка в Т-клетки еще до проникновения в тимус. Хотя стволовые клетки экспрессируют CD7, многие данные указывают на их полипотентность. Из гемопоэтических клеток-предшественников, выделенных из тимуса, in vitro развиваются гранулоциты, АПК, НК, В-клетки и клетки миелоидного ряда. Это означает, что проникающие в зачаток тимуса костномозговые клетки сохраняют исходную полипотентность.
Созревание Т-клеток происходит по мере перемещения тимоцитов из корковой зоны в мозговую. Тимус состоит из долек, в каждой из которых различают корковую и мозговую зоны (см. гл. 3). В этих зонах присутствуют эпителиальные клетки, макрофаги и имеющие костномозговое происхождение интерлигитатные клетки с высоким уровнем экспрессии антигенов МНС класса II. Для дифференцировки Т-лимфоцитов необходимы клетки всех этих трех типов (рис. 12.6). Например, специализированные эпителиальные клетки из периферических областей корковой зоны тимуса (клетки-«няни») содержат тимоциты в своих цитоплазматических «карманах» и могут участвовать в процессе их «обучения» (см. ниже). Поступающие из костного мозга стволовые клетки в первую очередь колонизируют подкапсульный слой тимуса. Они развиваются в крупные, активно пролиферирующие лимфобласты, которые и дают начало популяции тимоцитов.
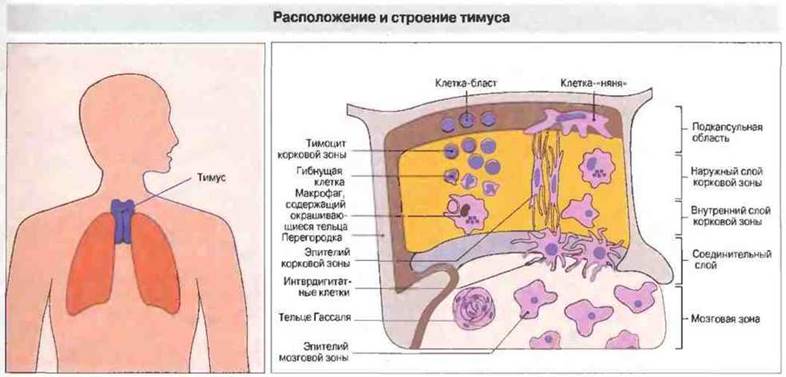
Рис. 12.6. Состоящий из двух долей тимус представляет собой орган, окруженный капсулой и разделенный перегородками на отдельные дольки. Корковая зона содержит плотно упакованные делящиеся корковые тимоциты в сети эпителиальных клеток, которые присутствуют и в мозговой зоне. Последняя содержит меньше лимфоцитов, но больше антигенпрезентирующих клеток костномозгового происхождения - интердигитатных клеток и макрофагов. Развивающиеся тимоциты тесно контактируют с эпителиальными и интердигитатными клетками, которых особенно много на границе корковой и мозговой зон. Функция извитых эпителиальных структур, называемых тельцами Гассаля, неизвестна.
В корковой зоне тимуса присутствует гораздо больше развивающихся лимфоцитов (85—90 %), чем в мозговой зоне. Изучение функции клеток и их поверхностных маркеров показывает, что тимоциты корковой зоны являются менее зрелыми, чем тимоциты мозговой зоны. Судя по этому, тимоциты мигрируют из коркового слоя в мозговой, где происходит их созревание. Полностью созревшие Т-клетки покидают тимус через посткапиллярные венулы, расположенные в зоне соединения коркового и мозгового слоев. Однако могут существовать и другие пути выхода клеток из тимуса, в том числе через лимфатические сосуды.
В процессе созревания Т-клетки меняют свой фенотип. Процесс превращения стволовых клеток в зрелые Т-клетки, как и созревание гранулоцитов и моноцитов, сопровождается появлением или исчезновением на их поверхности «дифференцировочных» маркеров, имеющих функциональное значение. Анализ генов, кодирующих αβ- и γδ-рецепторы Т-клеток, а также изучение смены поверхностных антигенов показывают, что дифференцировка Т-клеток в тимусе происходит по меньшей мере в двух направлениях. Неясно, различаются ли эти пути с самого начала; вероятнее всего, они представляют собой ответвления от одного общего исходного пути. Лишь очень небольшая доля зрелых лимфоцитов тимуса (менее 1 %) экспрессирует γδ-ТкР. Большинство же тимоцитов дифференцируется в клетки с αβ-ТкР; на их долю приходится более 99 % Т-лимфоцитов, присутствующих во вторичных лимфоидных тканях и крови (см. гл. 2 и 3).
Фенотипический анализ обнаруживает последовательные изменения в антигенном составе клеточной мембраны при созревании Т-клеток (рис. 12.7). Изменения фенотипа упрощенно можно представить в виде трехстадийной модели.
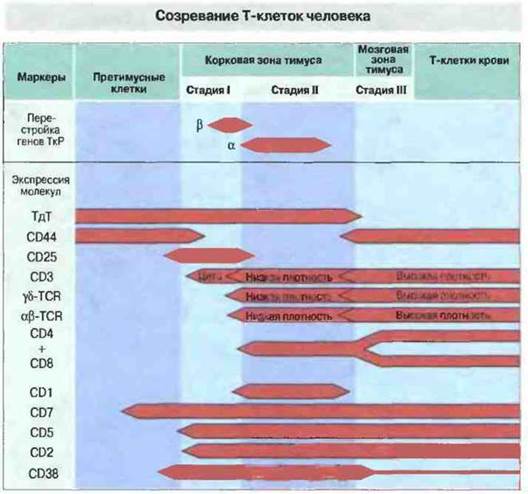
Рис. 12.7. Экспрессия Т-клеточных маркеров в процессе созревания Т-клеток у человека. В стволовых клетках тимуса присутствует фермент терминальная дезоксинуклеотидил-трансфераза (ТдТ); на стадии II ее активность уменьшается, а в клетках мозговой зоны полностью исчезает. В процессе дифференцировки появляется несколько поверхностных гликопротеинов. На мебране корковых тимоцитов, находящихся на II стадии развития, присутствует молекула CD1, которой уже нет на клетках мозгового слоя. Маркеры CD2 и CD7 (общий маркер Т-клеток) появляются на очень ранних стадиях дифференцировки и сохраняются до стадии зрелой Т-клетки. Зрелые Т-клетки сохраняют и молекулу CD5, которая также появляется на ранней стадии. Молекула CD3 вначале, на стадии I, экспрессируется в цитоплазме (цито), а затем и на клеточной поверхности, одновременно с ТкР. У большинства клеток на стадии II поверхностные молекулы CD3 и αβ-ТкР присутствуют в небольшой концентрации, но она значительно возрастает на стадии III. На стадии II одновременно экспрессируются молекулы CD4 и CD8, т. е. клетки дважды положительные. Одна из этих молекул при переходе клеток в III стадию (зрелые Т-клетки) теряется (моноположительные клетки).
Тимоциты I стадии (ранние). Стадия 1 включает две фазы. В первой фазе клетки экспрессируют CD44 и CD25, но при этом они дважды отрицательные — CD4-, CD8-; гены ТкР сохраняют гаметную конфигурацию. Клетки, находящиеся в этой фазе, способны дифференцироваться и в других направлениях (см. выше). Во второй фазе они теряют CD44, но все еще остаются отрицательными и по CD4, и по CD8; перестраивается ген β-цепи ТкР. В этот период тимоциты экспрессируют цитоплазматическую (но не мембраносвязанную) форму молекулы CD3, образующей комплекс с ТкР, и таким образом коммитированы к дифференцировке в Т-клетки. Экспрессия CD7, наряду с CD2 и CD5, продолжается. На этой стадии экспрессируются и маркеры пролиферации, такие как рецептор трасферрина (CD71) и CD38 (общий маркер всех ранних предшественников гемопоэтических клеток). Следует обратить внимание на то, что ни один из маркеров пролиферации не специфичен для Т-клеточного пути дифференцировки. Однако для ранних тимоцитов этот путь предопределяется перестройкой гена β-цепи ТкР и экспрессией в цитоплазме (но не на мембране) комплекса CD3.
Тимоциты II стадии (промежуточные, или часто встречающиеся). На долю этих клеток всегда приходится примерно 85 % всех лимфоидных клеток тимуса. Для них характерен фенотип CD1+, CD44-, CD25-, но при этом они дважды положительные — CD4+, CD8+. В промежуточных тимоцитах происходит перестройка генов, кодирующих α-цепь ТкР; на клеточной поверхности с низкой плотностью экспрессируются обе цепи αβ-рецептора в ассоциации с комплексом CD3.
Тимоциты III стадии (зрелые). На этой стадии происходят резкие изменения фенотипа клеток, а именно потеря CD1, экспрессия на мембране с высокой плотностью комплекса αβ-ТкР - CD3 и разделение клеток на два подтипа, экспрессирующих один CD4, другой CD8 (т. е. клетки становятся моноположительными). Большинство тимоцитов на этой стадии лишены CD38 и рецептора трансферрина и их практически невозможно отличить от зрелых Т-клеток крови. Все эти клетки, обнаруживаемые в мозговой зоне тимуса, экспрессируют рецептор CD44, предположительно участвующий в миграции и хоминге лимфоцитов в периферических лимфоидных тканях. На этой стадии экспрессируется также L-селектин.
Разнообразие T-клеточных рецепторов формируется в тимусе. Т-клетки способны распознавать огромное количество разнообразных антигенов. В процессе созревания этих клеток в тимусе гены αβ- и γδ-ТкР претерпевают соматическую рекомбинацию, образуя функциональные гены для различных Т-клеточных рецепторов (см. гл. 8). Цепи β и δ кодируются сегментами V, D и J, тогда как для синтеза α- и γ-цепей служат только сегменты V и J. Первыми в процессе созревания Т-клеток перестраиваются гены ТкР, кодирующие γ-цепи, а затем уже гены β- и α-цепей. В результате случайных сочетаний разных генных сегментов возникает множество продуктивных перестроек. Это обеспечивает экспрессию разнообразных пептидных последовательностей вариабельных участков обеих цепей ТкР. Тимоциты, в которых перестройка генов оказывается непродуктивной, погибают. Как и при создании разнообразия В-клеточных рецепторов (антител), важнейшую роль в процессе перестройки, обусловливающей разнообразие Т-клеточных рецепторов для антигенов, играют два активирующих рекомбинацию гена — RAG-1 и RAG-2 (см. гл. 7).
Вначале ТкР экспрессируются на клеточной поверхности с низкой плотностью. Это характерно для Т клеток под капсульного и наружного слоев корковой зоны тимуса, в которых клетки активно пролиферируют.
«Альтернативные» формы ТкР в процессе созревания. Исследования на трансгенных мышах показали, что в ранней стадии онтогенеза Т-клетки могут экспрессировать альтернативные формы ТкР, которые, возможно, участвуют в передаче дифференцировочных сигналов. Это димеры β-ТкР, ассоциированные с CD3 в отсутствие α-ТкР; мембраносвязанные цепи β-ТкР, ассоциированные с фосфатидилинозитолом, а не CD3; β-цепи ТкР, ассоциированные на поверхности клетки с неполным комплексом CD3 (CD3 лишь из γ-, β-, и ε- но без ζ- и η-цепей) и без α-цепи ТкР. Наконец, возможна экспрессия «суррогатной» α-цепи (пре-Т α-цепь), роль которой, по-видимому, выполняет недавно идентифицированный гликопротеин 33 кДа. Не исключено, что такие рецепторы, как и «суррогатные» пре-В-клеточные рецепторы (см. ниже), принимают участие в процессах пролиферации, созревания и селекции на ранних стадиях дифференцировки лимфоцитов.
В тимусе происходит положительная и отрицательная селекция развивающихся Т-клеток
Положительная селекция. Т-клетки распознают антигенные пептиды только представленными в «контексте» собственных молекул МНС на поверхности АПК (см. гл. 7 и 9). В действительности Т-клетки осуществляют двойное распознавание - и антигенных пептидов, и полиморфной части молекул МНС. (Маркер CD4, присутствующий на поверхности Т-клеток одной из субпопуляций, распознает неполиморфную часть молекул класса II.) Положительная селекция (называемая также «обучением в тимусе») заключается в том, что дальнейшей дифференцировке подвергаются только те клетки, ТкР которых обладают невысокой аффинностью к собственным молекулам МНС. По имеющимся данным, положительную селекцию осуществляют эпителиальные клетки тимуса, выступающие в роли АПК (рис. 12.8). Т-клетки, рецепторы которых обладают очень высокой или очень низкой аффинностью к собственным молекулам МНС, подвергаются в корковой зоне тимуса апоптозу и погибают. Апоптоз — это запрограммированное «самоубийство» клетки, осуществляемое активированными эндогенными нуклеазами путем расщепления ДНК на фрагменты (рис. 12.9).
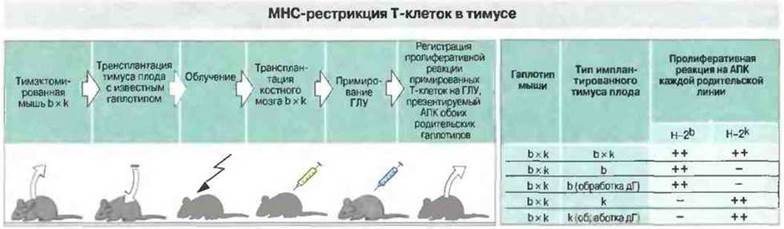
Рис. 12.8. Тимэктомированным мышам [F1 (Н-2b х Н-2k] имплантировали тимусы 12-суточных эмбрионов различных генотипов. Затем мышей облучали дли элиминации собственных Т-клеток и пересаживали им костный мозг от особей F1, обеспечивающий организм стволовыми клетками. После сенсибилизации антигеном (гемоцианин лимфы улитки, ГЛУ) регистрировали пролиферативные реакции Т-клеток лимфоузлов на ГЛУ, презентируемый АПК особей каждой родительской линии. В некоторых опытах доли тимуса перед имплантацией инкубировали с дезоксигуанозином (дГ), который разрушает внутритимусные макрофаги и дендритные клетки. Результаты показали: 1) для того, чтобы Т-клетки «научились» распознавать молекулы МНС, необходимо соответствующее тимусное окружение и 2) макрофаги и дендритные клетки костномозгового происхождения (удаляемые при обработке дГ) для этого не нужны. (По Lo D., Sprent V. 1986. Nature 319: 672.)
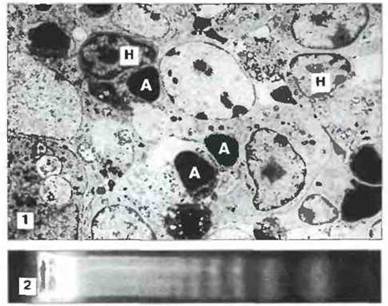
Рис. 12.9. Апоптоз клеток в тимусе. 1. К долькам тимуса плода в тканевой культуре добавляли антитела анти-СD3, в результате чего при участии ТкР происходила активация клеток и включался механизм их запрограммированной гибели (апоптоз). На электронной микрофотографии видно, что в отличие от ядер нормальных клеток (Н), в которых хроматин диспергирован, в ядрах апоптотических клеток (А) наблюдается выраженная конденсация хроматина. (Фото любезно предоставлено д-ром С Smith.) 2. Анализ ДНК апоптотических клеток методом электрофореза в агарозном геле; видна характерная «лесенка» из фрагментов расщепленной ДНК.
Т-клетки с рецепторами, обладающими невысокой аффинностью, избегают апоптоза, выживают и продолжают путь созревания.
Отрицательная селекция. Некоторые Т-клетки, прошедшие положительную селекцию, могут обладать рецепторами, распознающими не молекулы МНС, а другие компоненты собственных тканей. Такие клетки выбраковываются путем «отрицательной селекции», происходящей в более глубоких слоях корковой зоны тимуса, в месте соединения корковой и мозговой зон и в мозговой зоне. Тимоциты взаимодействуют с собственными антигенами, которые презентируются интердигитатными клетками. Дальнейшее созревание «разрешается» только тем тимоцитам, которые лишены способности распознавать собственные антигены; остальные подвергаются апоптозу и разрушаются, Эти отмирающие тимоциты, как и любые другие апоптотические клетки тимуса, в глубоких слоях корковой зоны фагоцитируются макрофагами, содержащими окрашивающиеся тельца. Существование отрицательной селекции (называемой также центральной толерантностью) недавно было убедительно доказано в исследованиях на мышах, у которых экспрессированные в тимусе эндогенные суперантигены вызывают элиминацию Т-клеток, несущих ТкР с той или иной Vβ-цепью (см. гл. 13).
Т-клетки на этой стадии созревания (CD4+, CD8+, ТкРlo) продолжают экспрессировать ТкР с высокой плотностью, но теряют либо CD4, либо CD8, становясь моноположительными зрелыми тимоцитами. Эти разные субпопуляции CD4+- и СD8+-клеток, обладая специальными рецепторами хоминга (например, CD44), мигрируют в периферические лимфоидные ткани, где функционируют как зрелые хелперные и цитотоксические Т-клетки соответственно. Тимус покидает менее 5 % тимоцитов; остальные погибают в процессе селекции (рис. 12.10) или вследствие неспособности экспрессировать антигенные рецепторы.
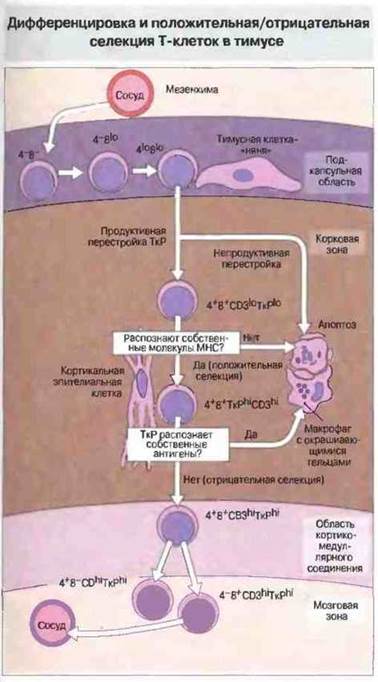
Рис. 12.10. Согласно этой гипотетической схеме, «претимические» Т-клетки направляются в закладку тимуса и проникают в нее. В подкапсульном слое они пролиферируют, образуя крупные лимфобласты, которые, размножаясь, дают начало популяции клеток, вступающих на путь дифференцировки. Многие из этих клеток находятся в тесном контакте с эпителиальными клетками-«нянями» тимуса (значение такого взаимодействия остается спорным вопросом). В этом слое клетки впервые начинают экспрессировать CD8, а затем и CD4, оба с низкой плотностью. В них происходит также перестройка генов ТкР и может начаться экспрессия в небольшом количестве продуктов этих генов на поверхности клеток. Созревающие клетки перемещаются в более глубокие слои корковой зоны и контактируют здесь с эпителиальными клетками. Последние удлиняются и ветвятся, создавая тем самым большую поверхность для контакта с тимоцитами. Этот контакт обеспечивает «обзор» ТкР тимоцитов молекулами МНС, экспрессированными на поверхности эпителиальных клеток. В результате происходит положительная селекция. Отбракованные клетки подвергаются апоптозу и поглощаются макрофагами. Во время миграции тимоцитов из подкапсульного слоя глубже в корковую зону в них усиливается экспрессия CD3, ТкР, CD4 и CD8. В зоне кортико-медуллярного соединения аутореактивные тимоциты, т. е. распознающие аутоантигены, презентируемые интердигитатными клетками и макрофагами, подвергаются делеции. Этот процесс носит название отрицательной селекции, и в нем могут принимать участие также эпителиальные клетки мозговой зоны тимуса. После этой стадии формируются клетки, экспрессирующие либо CD4, либо CD8. Они покидают тимус через специальные сосуды соединительного слоя. (Процесс отрицательной селекции может происходить и в корковой зоне тимуса, приводя к элиминации тех клеток, ТкР которых обладают высокой аффинностью к собственным молекулам МНС.)
Роль молекул адгезии и цитокинов в созревании тимоцитов. Важнейший момент в дифференцировке Т-клеток это адгезия (прилипание) созревающих тимоцитов к эпителиальным и вспомогательным клеткам тимуса. Она происходит за счет взаимодействия комплементарных молекул адгезии, например CD2 с LFA-3 (CD58) и LFA-1 (CD11a, CD18) с ICAM-1 (CD54).
В результате этого взаимодействия индуцируется синтез цитокинов ИЛ-1, ИЛ-3, ИЛ-6 и ГМ-КСФ, необходимых для созревания Т-клеток. На ранних стадиях созревания тимоциты экспрессируют также рецептор для ИЛ-2. Этот цитокин вместе с другими молекулами способствует пролиферации клеток, которая происходит главным образом в подкапсульном слое и наружных слоях корковой зоны тимуса.
Некоторые Т-клетки созревают вне тимуса
Отрицательная селекция может осуществляться в периферических лимфоидных тканях. При созревании в тимусе элиминируются не все аутореактивные Т-клетки. Это, по-видимому, связано с тем, что не все аутоантигены способны проходить через тимус. Эпителиальный 6apьep тимуса может ограничивать также доступность некоторых антигенов из крови. Поскольку часть аутореактивных Т-клеток выживает, для предотвращения их реакции на собственные ткани организма необходим дополнительный механизм. Недавно проведенные эксперименты на трансгенных мышах позволяют предполагать, что для периферической инактивации аутореактивных Т-клеток (периферическая толерантность) может существовать два механизма:
✵ подавление (понижающая регуляция) экспрессии ТкР и CD8 (на цитотоксических клетках), вследствие которой такие клетки теряют способность взаимодействовать с аутоантигенами-мишенями, и
✵ анергия, развивающаяся при отсутствии необходимых вторых сигналов активации, источником которых служат клетки-мишени (например, при участии CD28).
Периферическая толерантность подробнее рассмотрена в гл. 14.
Созревание Т-клеток вне тимуса. Хотя для дифференцировки подавляющего большинства Т-клеток необходим функционирующий тимус, небольшое количество клеток (часто олигоклональной природы), обладающих Т-клеточными маркерами, обнаруживается и у бестимусных (nude) мышей. Не исключено присутствие у таких мышей закладки тимуса, однако все больше данных указывает на то, что костномозговые предшественники могут заселять эпителий слизистых оболочек, созревая там без участия тимуса в функциональные Т-клетки, экспрессирующие у5-ТкР и, возможно, также αβ-ТкР. Значение внетимусного созревания Т-клеток у эутимических животных (т. е. имеющих нормальный тимус) пока остается неясным.
Т-клетки новорожденных неполностью зрелые
Большинство Т-клеток, присутствующих в крови новорожденных, несут маркер CD45RA; это свидетельствует, что они еще не встречались с антигеном. Кроме того, при взаимодействии с различными антигенами Т-клетки новорожденных вырабатывают меньше интерферона-γ (и, вероятно, других цитокинов), чем зрелые Т-клетки.
В-клетки млекопитающих созревают в костном мозге и печени плода
У млекопитающих специальный орган для лимфопоэза В-клеток отсутствует. Эти клетки развиваются непосредственно из лимфоидных стволовых клеток в гемопоэтической ткани в печени плода (рис. 12.11), у человека на 8-9 неделе, у мыши примерно на 14 сутки внутриутробного развития. Позднее образование В-клеток происходит уже не в печени, а в костном мозге, где и продолжается в течение всей жизни организма. То же относится и к другим направлениям дифференцировки гемопоэтических клеток — эритроцитарному, гранулоцитарному, моноцитарному и тромбоцитарному. Недавно показано, что у мыши и человека во время внутриутробного развития предшественники В-клеток присутствуют также в ткани сальника. Появляются ли они здесь раньше, чем в закладке печеночной ткани плода, неизвестно.
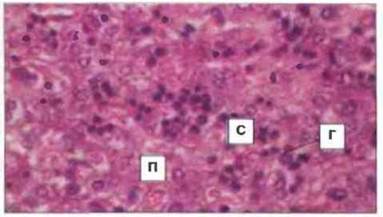
Рис. 12.11. Срез печени плода человека. Видны островки гемопоэза (Г); гемопоэтические стволовые клетки обнаруживаются в синусоидальных пространствах (С) между пластами печеночных клеток (П). (Фото любезно предоставлено д-ром A. Stevens и проф. J. Lowe.)
Образование В-клеток в костном мозге происходит не в отдельных участках. В-клетки-предшественники в костном мозге примыкают к эндосту костной пластинки. Каждая В-клетка-предшественник на стадии перестройки генов иммуноглобулинов может давать до 64 клеток-потомков, и эти клетки мигрируют к центру каждой полости губчатой кости, достигая просвета венозного синусоида. Созревание В-клеток в костном мозге происходит при их тесном контакте с клетками стромы, расположенными как вблизи эндоста, так и в окружении центрального синуса, где они называются адвентициальными клетками (рис. 12.12). Ретикулярные клетки имеют смешанные фенотипические признаки, будучи сходны по некоторым из них с фибробластами, эндотелиальными клетками и миофибробластами. Они продуцируют коллаген IV типа, ламинин и гладкомышечную форму актина. Эксперименты in vitro показывают, что стромальные клетки поддерживают дифференцировку В-клеток, возможно за счет продукции ИЛ-7. Адвентициальные клетки могут играть важную роль в процессе высвобождения зрелых В-клеток в центральный синус.
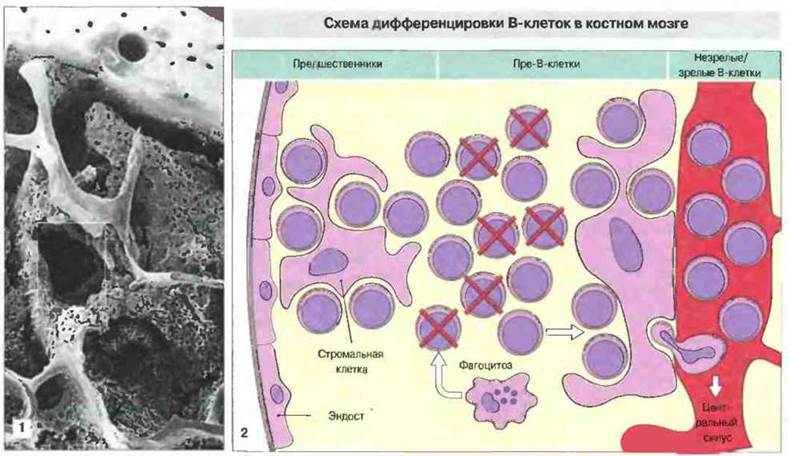
Рис. 12.12. 1. Микрофотография, полученная методом сканирующей электронной микроскопии низкого напряжения; видны строение кости и ее связь с костным мозгом. Выделенная квадратом полость схематически изображена справа. (Фото любезно предоставлено д-ром A. Stevens и проф. J. Lowe.) 2. Созревание В-клеток в полостях губчатой кости происходит по мере их перемещения в радиальном направлении к центру (от эндоста к центральному венозному синусу). Незрелые клетки-предшественники, примыкающие к слою клеток эндоста, дифференцируются в пре-В-клетки, многие из которых погибают и поглощаются костномозговыми макрофагами, содержащими окрашивающиеся тельца. Выжившие клетки продолжают созревать и достигают центрального венозного синуса. На всех стадиях созревания В-клеток важную роль играют связь с клетками стромы и присутствие цитокинов, таких как ИЛ-7. (По Osmond D., Gallagher R. 1991. Immunol. Today 12: 1-3.)
В-клетки подвергаются процессам селекции. Большинство созревающих в костном мозге В-клеток (более 75%) не попадает в кровоток, а подобно тимоцитам погибает в результате апоптоза и поглощения костномозговыми макрофагами. Предполагается, что при взаимодействии В-клеток с клетками стромы происходит своего рода положительная селекция, которая «спасает» от запрограммированной гибели небольшую часть В-клеток с продуктивной перестройкой генов иммуноглобулинов. Отрицательная селекция аутореактивных В-клеток может происходить в костном мозге или селезенке — органе, в который мигрирует большинство новообразованных В-клеток в период внутриутробного развития.
Кинетические исследования позволяют рассчитать, что у мыши ежесуточно образуется примерно 5 · 107В -клеток. Поскольку селезенка мыши содержит приблизительно 7,5 · 107 В-клеток, огромная часть их должна погибать, что происходит, вероятно, на стадии пре-В-клеток из-за непродуктивной перестройки рецепторных генов или из-за экспрессии этими клетками аутореактивных иммуноглобулиновых рецепторов.
Маркерами зрелых В-клеток служат иммуноглобулины. Лимфоидные стволовые клетки, экспрессирующие терминальную дезоксинуклеотидилтрансферазу (ТдТ), пролиферируют, дифференцируются и претерпевают перестройку генов иммуноглобулинов (см. гл. 8), что приводит к образованию пре-В-клеток, в цитоплазме которых появляются тяжелые μ-цепи. Некоторые из этих пре-В-клеток и на своей поверхности несут небольшое количество μ-цепей, ассоциированных с «суррогатными» легкими цепями, Vnpe-В и λ5 (см. ниже; схема строения пре-В-клеточного рецептора приведена на рис. 12.13). К этому времени уже происходит аллельное исключение либо материнских, либо отцовских генов иммуноглобулинов. Из пролиферирующих крупных пре-В-клеток образуются пре-В-клетки меньших размеров. Как только В-клетка начинает синтезировать легкие цепи, которые могут быть k- или λ-типа, ее антигенный рецептор sIgMприобретает антигенсвязываюшую специфичность. Таким образом, одна В-клетка способна производить антитела лишь одной специфичности — основное положение теории клональной селекции относительно продукции антител. В стадии про-В-клеток на клеточной поверхности появляются ассоциированные с иммуноглобулинами молекулы Igα и Igβ (CD79a и b). Схема дифференцировки В-клеток, сопровождающейся экспрессией иммуноглобулинов и некоторых других важных молекул, приведена на рис. 12.14.
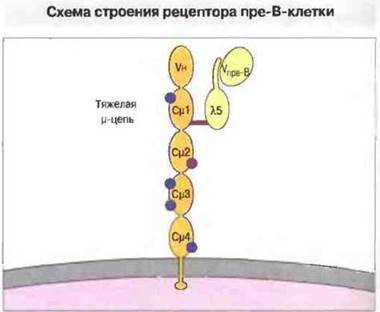
Рис. 12.13. Суррогатный В-клеточный рецептор состоит из тяжелой μ-цепи и белков Vnpe-B и λ5 (суррогатные легкие цепи), нековалентно связанных друг с другом. Белок λ5 ковалентно связан с доменом Сμ1. Функция рецептора пре-В-клетки неизвестна; возможно, он (подобно гипотетическому рецептору пре-Т-клетки) принимает участие в процессах ранней дифференцировки, селекции и пролиферации. Синие кружки - участки гликозилирования.
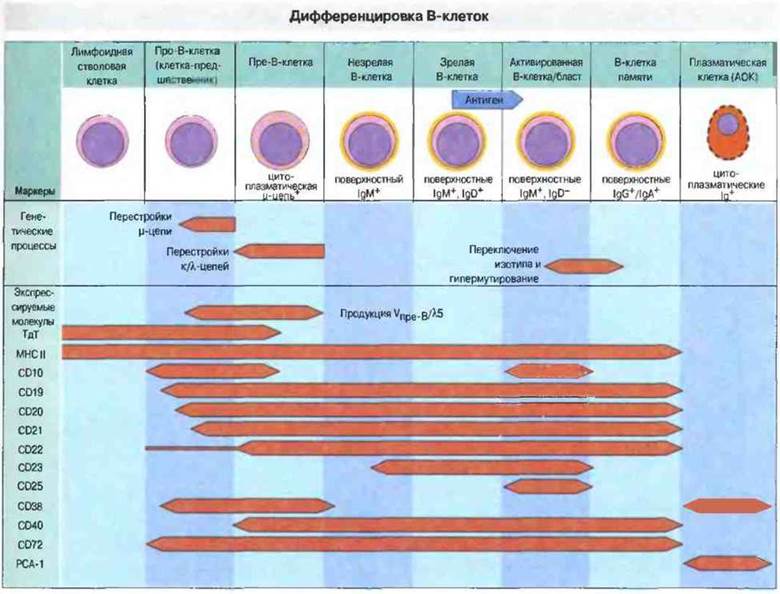
Рис. 12.14. Из лимфоидных стволовых клеток образуются девственные (нестимулированные) В-клетки, которые под действием антигена могут превращаться в клетки иммунологической памяти или в плазматические клетки. Желтым цветом показана локализация иммуноглобулинов в клетке. В процессе созревания клеток-предщественников происходит перестройка генов, кодирующих антитела. Пре-В-клетки экспрессируют только цитоплазматическую μ-цепь. Некоторые из них могут экспрессировать свои μ-цепи в ассоциации с суррогатными легкими цепями (рис. 12.13). Незрелые В-клетки несут на поверхности IgM, а зрелые - иммуноглобулины других иэотипов. При антигенной стимуляции В-клетки пролиферируют и после фазы пролиферации, активации и бласттрансформации превращаются либо в плазматические клетки, либо в клетки иммунологической памяти. Клетки памяти и плазматические клетки присутствуют в различных лимфоидных тканях. Терминальная дезоксинуклеотидилтрансфераза (ТдТ) экспрессируется на очень ранних стадиях онтогенеза. На схеме показана также последовательность появления других важных поверхностных маркеров В-клеток. Плазмоцитарный антиген-1 (РСА-1) обнаруживается только на плазматических клетках. Обратите внимание, что CD38 - это одна из молекул, которые присутствуют на поверхности ранних клеток-предшественников, затем исчезают и вновь появляются на мембране уже полностью дифференцированных плазматических клеток.
На развивающихся В-клетках появляются характерные молекулы поверхности. В процессе развития В-клеток происходят перестройка генов иммуноглобулинов и фенотипические изменения, сходные с описанными выше для Т-клеток. Самым ранним указанием на начало В-клеточной линии дифференцировки служит перестройка генов тяжелых цепей Ig в В-клетках- предшественниках. На последующих стадиях развития пре-В-клеток перестраиваются гены легких цепей. Раньше начала синтеза иммуноглобулинов экспрессируются некоторые поверхностные маркеры В-клеток, а именно молекулы МНС класса II, CD19, CD20, CD21, CD40 и антиген CD10 (CALLA). Последний из этих маркеров представляет собой высококонсервативную нейтральную эндопептидазу, временно экспрессируемую на ранних прешественниках В-клеток еще до появления в цитоплазме тяжелых μ-цепей. Позднее, уже после активации антигеном, В-клетки вновь начинают экспрессировать антиген CALLA (рис. 12.14). Другие маркеры, например CD23 и CD25 (а-рецептор для ИЛ-2), обнаруживаются главным образом на активированных В-клетках.
Ранние этапы развития В-клеток зависят от ряда факторов роста и дифференцировки. На разных стадиях дифференцировки В-клетки экспрессируют рецепторы для этих факторов. Процесс дифференцировки инициируют ИЛ-7, ИЛ-3 и низкомолекулярный фактор роста В-клеток (L-BCGF), тогда как на последующих стадиях действуют иные факторы (рис. 12.15).
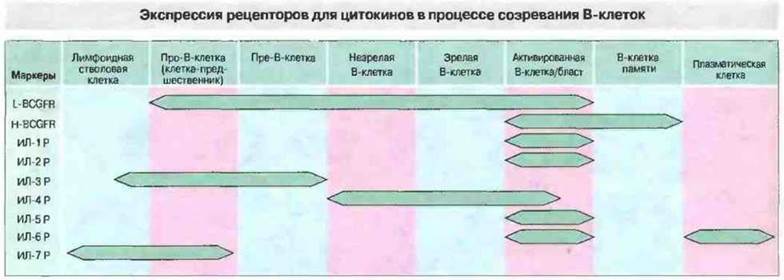
Рис. 12.15. Весь жизненным цикл В-клеток от стволовой до зрелой плазматической клетки регулируется присутствующими в их микроокружении цитокинами. (Важную роль в начале дифференцировки играет ИЛ-7.) Рецепторы для этих цитокинов избирательно экспрессируются В-клетками на различных стадиях дифференцировки. Некоторые из таких рецепторов теперь имеют обозначение как CD-маркеры (см. приложение).
В-клетки мигрируют во вторичные лимфоидные ткани, где осуществляют свои функции. Ранние В-клетки-«иммигранты» (у человека на 17 неделе внутриутробного развития) в селезенке и лимфоузлах плода являются sIgM+-клетками, на поверхности которых присутствует CD5 (предполагаемый маркер Т-клеток). В-клетки-предшественники CD5+ обнаруживаются также в сальнике плода и в зоне мантии вокруг вторичных фолликулов зрелых лимфоузлов.
После антигенной стимуляции зрелые В-клетки могут превращаться в клетки иммунологической памяти или в антителообразующие клетки (АОК). Плазматические клетки (окончательно дифференцированная форма АОК) обычно теряют поверхностные иммуноглобулины (sIg), поскольку функция этих Ig в качестве рецепторов им больше не нужна. Подобно всем другим окончательно дифференцированным гемопоэтическим клеткам плазматическая клетка имеет ограниченную продолжительность жизни и в конце концов подвергается апоптозу (см. рис. 2.23).
Незрелые и зрелые В-клетки отвечают на антиген по-разному. При обработке антителами анти-IgM или антигеном и те и другие теряют sIgM, молекулы которого удаляются путем кэппинга и эндоцитоза. Однако ресинтез sIgM наблюдается в культуре только у зрелых В-клеток (рис. 12.16). Такая индуцированная потеря антигенного рецептора может служить одним из механизмов приобретения аутореактивными В-клетками толерантности в процессе их созревания.
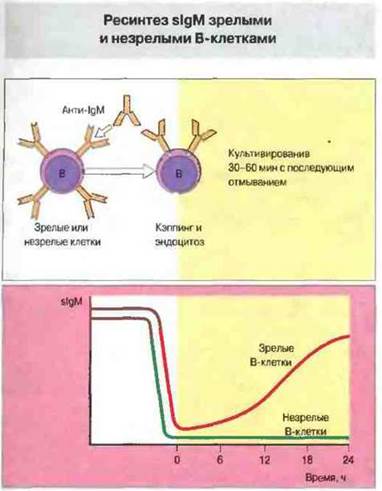
Рис. 12.16. Зрелые (от взрослых) и незрелые (от новорожденных) В-клетки инкубировали при 37 °С с антителами к sIgM (анти-IgM) в течение 30-60 мин; это приводило к кэппингу молекул sIgM и их интернализации путем эндоцитоза. Затем клетки отмывали от добавленных антител. Как видно на рисунке, только зрелые В-клетки за последующие 24 ч восстанавливали синтез sIgM.
У птиц В-клетки созревают в фабрициевой сумке
Лимфопоэз В-клеток у птиц начинается в специальном лимфоэпителиальном органе — фабрициевой сумке. Закладка ее развивается из выпячивания эндодермы задней кишки и заселяется стволовыми клетками из крови. Исследования на химерах курица/куропатка показывают, что стволовые клетки проникают в сумку только в период между 10 и 14 сутками эмбрионального развития (см. гл. 15). Пиронинофильные клетки (предположительно стволовые) находятся в тесном контакте с эпителиальными клетками. Пролиферирующие клетки сумки образуют корковый и мозговой слои каждого фолликула, который может заселяться одной или несколькими стволовыми клетками.