Эволюция вирусов - Жданов В. М. 1990
Специальная часть
Парвовирусы
Парвовирусы (Parvoviridae)—это большая неоднородная группа вирусов, в которую входят вирусы, вызывающие разнообразные заболевания млекопитающих и птиц, сателлиты (дефектные вирусы) аденовирусов, вирусы насекомых [Ананьев В. А., 1982; Bachman Р. et al., 1979; Matthews R., 1982]. Всех их объединяет то, что геномом является небольшая, длиной около 5000 нуклеотидов (1,5х106—2,0х106), однонитевая ДНК. Нередко в разных вирионах содержатся положительная и комплементарная ей нити ДНК. Поэтому при экстракции ее из вирионов может образовываться двунитевая структура.
Семейство парвовирусов включает 3 довольно разные группы вирусов: возбудители своеобразных заболеваний млекопитающих и птиц (род Parvovirus), дефектные сателлиты аденовирусов (род Dependovirus) и вирусы денсонуклеоза насекомых (род Densovirus). Несмотря на экологическую разобщенность этих 3 групп вирусов, они имеют много общего как в строении вирионов, так и стратегии их генома.
Вирионы являются икосаэдрами, имеют диаметр 18—26 нм, 32 капсомера; диаметр капсомеров 3—4 нм. Вирионы имеют высокую плотность (1,34—1,42 г/мл) в связи с большим (до 20% массы) содержанием в них ДНК и небольшой коэффициент седиментации (110—120S). Генетический материал представлен однонитевой линейной ДНК с молекулярной массой 1,5х106—1,8х106, состоящей из 4675—5176 нуклеотидов. Первые два рода различаются между собой тем, что у первого ДНК в вирионах всегда имеют положительную полярность, тогда как у второго в вирионах могут содержаться плюс или минус-нити. Кроме того, геномы этих двух групп, вирусов имеют и другие отличия, хотя Т-образные структуры, образуемые 3'-концевыми последовательностями обеих групп, чрезвычайно сходны. Сходна и структура двух рамок считывания (неструктурные и структурные белки) у вирусов сравниваемых групп.
Вирионы содержат три капсидных белка (VP1, VP2 и VP3), молекулярная масса которых варьирует у разных вирусов. По-видимому, синтез всех 3 структурных белков кодируется общей последовательностью генома и различия объясняются разными точками считывания и сплайсингом.
После адсорбции, проникновения в ядра клеток и депротеинизации начинает функционировать вирусный геном, транскрипция которого обеспечивается клеточной РНК-полимеразой, причем синтезируются белки (NS1, NS2 или только первый из них), участвующие в репликации вирусной ДНК. У сателлитов аденовирусов в этом процессе участвуют и белки аденовируса. Предполагают, что конец складывается, образуя шпильку, и функционирует как праймер синтеза ДНК, после чего происходят разрезание в области шпильки и разделение родительской или дочерней молекул ДНК. Механизмы транспортировки структурных белков с мест синтеза в цитоплазме в ядро изучены мало, как и сборка вирионов. К роду Parvovirus отнесены 14 вирусов, вызывающих разнообразные заболевания грызунов, домашних животных, норок, кроликов, хомяков, гусей. Вирионные белки имеют молекулярную массу 81 000—86000 (VP1), 64000—77 000 (VP2), 60 000—67000 (VP3). Вирусы реплицируются в ядрах интенсивно делящихся клеток. Впрочем, при изучении парвовируса панлейкопении кошек-было показано, что он может реплицироваться в клетках с остановленной (блокированной) фазой S [Lenghаus С. et al., 1985].
В репликации парвовирусов (Н1 вирус мышей) участвует некапсидный вирусспецифический белок NCVP1, который существует в двух формах — собственно NCVP1 (84 000) и NCVP1' (92 000). Оба белка появляются на ранней стадии инфекции [Ron D., Tal J., 1984]. Парвовирусы нередко вызывают латентную инфекцию, длительную персистенцию. По антигенной структуре выделяют 4 группы вирусов, имеющих внутри групп иммунологическую общность. При заражений клеток L лимфотропным парвовирусом мышей устанавливается персистенция, в ходе которой, с одной стороны, происходят мутации вируса, а с другой — мутации клеток, что типично для этого взаимоотношения двух партнеров и может быть определено как модель коэволюции [Ron D., Tal J., 1985].
Один из неструктурных белков парвовирусов весьма консервативен, в том числе у автономно реплицирующихся парвовирусов и сателлитов аденовирусов, а также у парвовируса человека (В19) [Shade R. et al., 1986].
Парвовирус мыши (MVM) имеет гены для синтеза двух неструктурных белков: NS1 (83 000) и NS2 (25000). Геном кодирует 3 транскрипционных единицы, которые совпадают, но обладают разными местами сплайсинга, в результате чего мРНК имеют молекулярную массу 48000, 3300 и 3000. РНК с массой 3000 кодирует синтез двух капсидных белков VP1 (83 000) и VP2 (64 000). Третий структурный белок VP3 (62 000) образуется после протеолитического выщепления NН2-концевого участка VP2 [Cotmore S., Tattersall Р., 1986].
При сравнении парвовируса собак с вирусом энтерита норок была выявлена 86% гомология их ДНК, что свидетельствует о близком родстве обоих вирусов [McMaster G. et al., 1981]. Парвовирус, вызывающий алеутскую болезнь норок (хроническое системное заболевание), имеет однонитевую ДНК с молекулярной массой 1,4x106, седиментирующую при 16S. Два вирусных пептида, закодированные на ней, имеют молекулярную массу 89 100 и 77 600 [Bloom М. et al., 1980].
Парвовирусы млекопитающих (кошек, норок и собак) имеют выраженное антигенное родство, хотя вызывают разные болезни: у кошек — панлейкопению. У норок — энцефалопатию, у собак — энтерит й миокардит [Surleraux М., Burtonbay G., 1984]. Последний вирус индуцирует синтез 3 полипептидов (67 000, 70 000 и 85 000), а также, возможно, еще одного полипептида (50 000). Автономный парвовирус человека был обнаружен у здоровых доноров, а также у лиц с кореобразной сыпью, суставными болями, лейкопенией и другими симптомами, включая вспышки инфекционной эритемы [Anderson М., Pattison J., 1984].
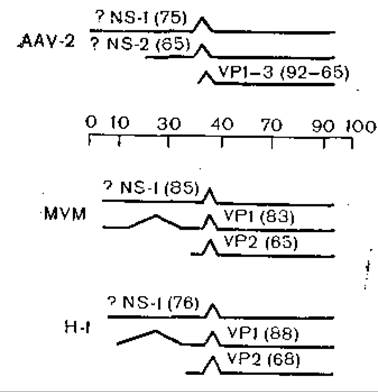
Рис. 40. Транскрипция ДНК-виру- сов AAV-2, MVM и Н-1. Горизонтальные линии — вирусные транскрипты, наклонные линии — нитроны; указаны области, кодирующие капсидные и неструктурные вирусные белки, в скобках — молекулярная масса белков (x103).
К роду Dependovirus отнесены сателлиты аденовирусов, выделенные вместе с аденовирусами человека, сельскохозяйственных животных и птиц. Молекулярная масса вирионных белков составляет 87 000 (VP1), 73 000 (VP2) и 62 000 (VP3).
Три капсидных белка содержат совпадающие аминокислотные последовательности. Их синтез кодируется на правой половине генома и для каждой из них синтезируется своя мРНК [Janik J. et al., 1984] (рис. 40).
Вирусы размножаются лишь в присутствии вируса-помощника, которым в естественных условиях является соответствующий аденовирус. Репликация сателлитов аденовирусов обеспечивается ранними генами аденовирусов (la, lb, 2а и 4), а также транскриптом малой РНК, синтезируемым клеточной РНК-полимеразой III [Richardson W., Westphal Н., 1984]. В эксперименте показана возможность размножения их в присутствии герпесвирусов [Buller R. et al., 1981].
При отсутствии аденовируса-хелпера репликации сателлита не происходит. Однако вирионы могут проникать в клетку, их ДНК может достичь ядра и там сохраниться, будучи интегрированной в клеточный геном. Освобождение ее может быть обеспечено вирусом-хелпером и стать началом продуктивной инфекции. Необходимо также отметить, что интеграция генома сателлита аденовируса не сопровождается трансформацией клетки и изменением ее фенотипа. В то время как аденовирус обеспечивает репликацию сателлита, последний при коинфекции угнетает как литический процесс, так и онкогенез, вызываемый аденовирусом. Вероятно, некоторые сателлиты аденовирусов, например ААВ4, могут поражать и человека, и домашних животных. Вся эта группа вирусов очень компактная и имеет общие «корни» происхождения.
Род Densovirus включает 10 вирусов чешуекрылых насекомых, а также, возможно, двукрылых и прямокрылых. В отличие от предыдущих родов представители этого рода имеют 4 вирионных белка: VP1 (18000), VP2 (72 000), VP3 (57 000) и VP4 (46 000). Вирусы размножаются преимущественно в личинках, накапливаясь в гипертрофированных ядрах.
Одними из интересных особенностей 4 структурных белков вируса денсонуклеоза тутового шелкопряда являются их выраженная гомология и наличие антигенного родства. По-видимому, синтез всех этих белков кодируется общей последовательностью ДНК. Гомология имеется также между разными видами денсовирусов насекомых [Bando Н. et al., 1984].
Репликация осуществляется при активном участии клеточной системы синтеза ДНК, поэтому вирусы развиваются преимущественно в делящихся клетках. При этом к ДНК ковалентно прикрепляется белок между тирозиновым остатком и 5'-концом молекулы ДНК. Этот белок не кодируется вирусом. [Chow М. et al., 1986].
Несмотря на сходство ряда свойств, парвовирусы все же не являются компактной группой обособленных вирусов, и объединение их в одно семейство может представляться несколько искусственным. Но, с другой стороны, эти вирусы и не являются случайной группировкой. Близкие размеры геномов, молекулярные массы структурных белков, тесная связь репликации с ядерной активностью клеток — все это объединяет вирусы теплокровных животных с вирусами насекомых.
Особенный интерес представляют дефектные вирусы. В данном случае это компактная группа сателлитов аденовирусов. Вообще же наряду с ДНК-содержащими имеются и РНК-содержащие вирусы, паразитирующие в растениях и грибах. Типичным вирусом-сателлитом, размножающимся в растениях, является сателлит вируса некроза табака. В данном случае типы нуклеиновой кислоты сателлита и вируса-помощника совпадают. В случае с дельта-вирусом дефектный вирус является РНК-содержащим, а реплицирующий его вирус — ДНК-содержащим. Однако необходима поправка: последний вирус имеет репликативным интермедиатом РНК. Происхождение этих многочисленных и разнообразных вирусов и их эволюция требуют специального изучения.