Эволюция вирусов - Жданов В. М. 1990
Общая часть
Основные направления эволюции биосферы
Цель этой главы — служебная: мы постараемся напомнить читателю основные факты и заключения, относящиеся к возникновению и эволюции органической жизни на Земле, остановившись, в частности, на эволюции групп животных и растений, которые стали хозяевами изученных вирусов.
В соответствии с современными представлениями возраст Земли определяется примерно в 4,7 (4,66) млрд лет, из которых около 1 млрд лет занимал догеологический, а 3,50— 3,57 млрд лет — теологический период. В свою очередь этот период делят на докембрий (3 млрд лет) и фенарозой (0,57— 0,585 млрд лет). Дальнейшее подразделение геологической истории Земли приведено в табл. 1.
Палеоген и неоген объединяют в третичный период и в этом случае антропогену соответствует четвертичный период. Но палеоген подразделяют на эпохи (палеоцен, эоцен и олигоцен), неоген подразделяют на миоцен и плиоцен, в этом случае антропогену соответствует плейстоцен (1—2 млн лет). Если считать временем возникновения Земли 4—5 млрд лет назад, то в начальном периоде, длившемся 0,5 млрд лет, поверхность Земли напоминала лунный рельеф. Затем Земля перешла в первично-океаническую стадию и образовалась первичная атмосфера. Тогда же, около 4 млрд лет назад, появилась кора континентального типа. К середине и особенно к концу архейской эры (3—2,5 млрд лет назад) прото континенты объединились в крупные плиты, а в начале мезозоя (1,4 млрд лет назад) образовался единый материк Пангея, состоявший из северной (Лавразия) и южной (Гендвана) частей. Затем, однако, произошли распад Гендваны и Лавразии (0,3 млрд лет назад) и формирование ныне существующих материков и океанов. Процесс этот был длительным, и до эоценового периода Австралия и Антарктида представляли еще единый материк. С конца эоцена (50 млн лет назад) конфигурация континентов и океанов приобретает современный вид.
Таблица 1. Геологическая хронология Земли
Эра |
Период |
Эпоха |
Возраст, млн лет с начала |
Продолжительность, млн лет |
Архей |
3570—3500 |
2000 |
||
Протерозой |
1570—1500 |
1000 |
||
Палеозой |
Кембрий |
585—570 |
50—80 |
|
Ордовик |
520—505 |
30—80 |
||
Силур |
490—425 |
50—70 |
||
Девон |
425—375 |
50—20 |
||
Карбон |
330—325 |
60—85 |
||
Пермь |
270—280 |
15—40 |
||
Мезозой |
Триас |
230—225 |
55—60 |
|
Юра |
175—165 |
30—45 |
||
Мел |
135—110 |
40—60 |
||
Кайнозой |
Третичный: |
75—70 |
68—73 |
|
палеоген |
75—70 |
40—55 |
||
Палеоцен |
75—70 |
10—15 |
||
Эоцен |
65—60 |
20—25 |
||
Олигоцен |
45—40 |
10—15 |
||
неоген |
35—25 |
24—33 |
||
Мноцен |
35—25 |
15—20 |
||
Плиоцен |
25—20 |
19—23 |
||
Четвертичный |
||||
Плейстоцен |
1,025—2 |
1—2 |
||
Плейстоцен |
1—2 |
1—2 |
||
Современная |
0,025 |
0,025 |
В истории Земли было несколько интенсивных периодов горообразования, отразившихся на эволюции биосферы. Первое великое горообразование произошло между археем и протерозоем, второе — на стыке протерозоя и кембрия. Оба они сопровождались значительным уничтожением ископаемых. В последующем были менее интенсивные горообразования, которые не сопровождались большим уничтожением ископаемых: герцинское — в конце перми и начале триаса, альпийское — в конце мезозоя, в неогене — в Северной Америке.
Первичные океаны составляли не более 1/10 объема нынешних, остальная их масса образовалась позже за счет дегазации внутренних частей Земли. Объем их значительно увеличился в докембрии и достиг современных размеров в кембрии. Соответственно менялись рельеф и климат Земли.
Архей характеризуется значительной вулканической деятельностью, эрозией на больших площадях и слабым процессом образования осадков. На грани архея и протерозоя произошел великий период горообразования. В это время очертания материков мало напоминали нынешние. В западном полушарии Канада и Гренландия образовали большой материк; Бразилия и Патагония были отдельными островами. Африка была связана с Аравией и Индией; большая часть Европы была покрыта морем. Восточная Европа образовывала материк Русской платформы, отделенный от островов Азии Уральским проливом и морем Течас. В протерозое, особенно в его конце, отмечаются выраженная вулканическая деятельность, многократные оледенения, а также интенсивный процесс образования осадочных пород.
В кембрии, после интенсивного горообразования, наряду с высокими горами на материках преобладал низменный рельеф, а климат был умеренным. Произошло перераспределение воды и суши. Канадско-Гренландский материк сначала превращается в архипелаг, а затем глубоко погружается. В южном полушарии Южная Америка существует как единый материк, включая Мексику, Флориду и Карибскую сушу. Африка соединена с Индией, Тибетом и Сунгаро-Гобией, образуя единый материк. В Европе остается большой островной массив Финко-Сарматии. Северо-восток Азии и северо-запад Америки образуют материк Берингию. Сибирское море меняет свои очертания, выходя в Атлантику и Пацифику, а в конце кембрия превращаясь в Центрально-Азиатский пролив.
В ордовике происходило значительное погружение суши и климат стал теплым даже в Антарктике. В силуре в связи с поднятием суши образовались обширные внутриконтинентальные моря, а климат стал более засушливым. В конце силура начинается каледонский горообразовательный процесс, приведший к возникновению ныне существующих Скандинавских гор, гор Казахстана, Саяно-Байкальских гор, гор Шотландии.
В девоне продолжается поднятие суши, внутриконтинентальные моря обмелевают и уменьшаются, климат становится аридным, а затем наступает оледенение. Появляются пустыни и полупустыни. В карбоне снова преобладают низменные материки, климат влажный и теплый, образуются обширные болота. В конце карбона вновь происходят поднятие суши и иссушение. В перми в связи с горообразованием материки приподняты, усиливается аридность, а в южном полушарии происходит оледенение.
В триасе продолжается развитие аридного климата, многие моря становятся мелководными, расширяются пустынные ландшафты. Эти же процессы продолжаются в юре, однако площадь тепловодных морей становится больше. В меловом периоде образуются многие горные системы в Европе, Азии и обеих Америках (Альпы, Гималаи, Скалистые горы, Анды). Климат становится более континентальным и прохладным. Сокращается площадь морей. В течение всего мезозоя образуются обильные отложения осадочных горных пород на всех континентах.
В кайнозое климат теплеет и остается таким в течение всего палеогена. В неогене холодает, а в течение плейстоцена были 4 периода оледенения, сменявшихся периодами потепления. Последний ледниковый период окончился 25 000 лет назад. Образовались, а затем исчезли сухопутные мосты между Европой и Англией, Азией и Северной Америкой, Индокитаем и Западным архипелагом. Материки приняли современное очертание.
Таковы были геологические и климатические условия, в которых происходили образование и эволюция органического мира на Земле.
Современные представления о происхождении жизни на Земле в значительной степени сложились под влиянием концепции А. И. Опарина (1957, 1966, 1968, 1980) и J. Haldane (1965). Отсылая читателя к этим работам, а также к трудам F. Crick (1968), М. Eigen (1971), М. Retten (1971) и других авторов, мы лишь напомним, что в соответствии с этими представлениями биологической эволюции предшествовала химическая эволюция, длившаяся более 1 млрд лет и приведшая к образованию первичных форм жизни на Земле.
В происхождении и эволюции жизни выделяют три главные стадии: преджизнь, раннюю жизнь в условиях бескислородной атмосферы и позднюю жизнь, связанную с кислородной атмосферой. Первичная атмосфера Земли была водородной, она исчезла в догеологическом периоде. В результате вулканической деятельности и процессов дегазации недр Земли к началу архейской эры образовалась вторичная атмосфера. Эта атмосфера была преимущественно восстановительной и содержала водород, азот, метан, аммиак, окись углерода и воду, а также цианид водорода. Более современная точка зрения заключается в том, что состав первичной атмосферы был не восстановительный, а нитрильный (3,5 Gyr). Он включал СО2, Н2О, N2и СО и меньше NH3, Н2, СН4 и Н2. При отсутствии кислородно-озонового экрана, под влиянием электрических разрядов и ультрафиолетового облучения стало возможным образование более сложных соединений: органических кислот, аминокислот, пуриновых и пиримидиновых оснований, порфиринов [Calvin М., 1969]. Все эти вещества, образовавшиеся в атмосфере, растворялись в водах Мирового океана, постепенно насыщая их и образуя «первичный бульон». За миллионы лет этот бульон стал достаточно густым, чтобы дать начало образованию высокополимерных органических соединений.
СХЕМА 1. ВОЗМОЖНЫЕ СОБЫТИЯ В ЭВОЛЮЦИИ ОТ «ПЕРВИЧНОГО БУЛЬОНА» К МНОГОКЛЕТОЧНОМУ ОРГАНИЗМУ

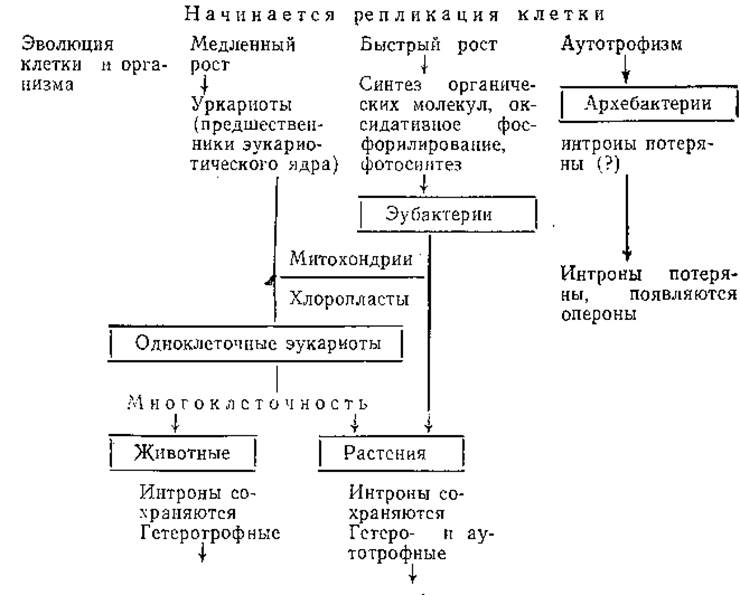
Накоплению продуктов «первичного бульона» способствовали орогенетические циклы развития Земли — чередование геосинклинальных орогенных и посторогенных периодов. Вследствие колебаний уровней моря в местах подъема суши в мелководных бассейнах под действием испарения могла происходить концентрация органических веществ «первичного бульона». Все это ускоряло химические реакции и способствовало накоплению первичных форм жизни и ее предшественников (схема 1) [Darnell A., Doolittle К., 1986].
В экспериментах, моделирующих эти первоначальные условия нашей планеты, были получены не только аминокислоты и азотистые основания, но и их полимеры — полипептиды и полинуклеотиды. По данным S. Fox и К. Dose (1977), в этих условиях образуются протеиноиды (протобелки), включающие все 20 аминокислот с молекулярными массами порядка нескольких тысяч, которые могут обладать каталитическими свойствами.
По сравнению с современной жизнью преджизнь обладала большим химическим разнообразием. Существенной ее основой явилась комплементариость пуриновых и пиримидиновых оснований, которые наряду с простыми аминокислотами (глицин, аланин, серин и пр.) образовались в первобытной атмосфере и океанах из паров аммиака, синильной кислоты, метана и двуокиси углерода. Образование полинуклеотидных цепей из пуриновых и пиримидиновых оснований создавало возможность их саморепликации с воспроизведением комплементарных полинуклеотидных цепей.
Дальнейший прогресс молекулярного отбора происходил при помощи аутокатализа, в результате которого возникли более сложные молекулы типа полипептидов, нуклеиновых кислот, полисахаридов. Вначале эти процессы были независимыми, но на каком-то этапе химической эволюции независимые процессы синтеза линейных последовательностей полипептидов и нуклеиновых кислот стали взаимосвязанными, поскольку такая объединенная система катализа имела преимущество перед двумя независимыми системами в том, что скорость реакции повышалась, а продукты реакции воспроизводились с высокой точностью. Таким образом мог возникнуть первичный генетический код. Вероятно, первичный генетический код был двоичным, и отражением его является решающее значение двух первых оснований современного трехчленного генетического кода. Вначале число аминокислот, из которых формировались полипептидные цепи, не превышало 10.
Такой примитивный генетический код сформировался более 3,5 млрд лет назад, и в это время появились протобионты — примитивные клеточные структуры с метаболизмом. Позже число аминокислот увеличивалось до 20, а генетический код стал трехбуквенным и избыточным (вырожденным). Универсальный генетический код формировался 3,5—2,5 млрд лет назад, и в это время появились палеокариоты с универсальным дизайном биохимических структур и их метаболизмом. Менее 2,5 млрд лет назад генетический код стал универсальным, появились прокариоты, а затем эукариоты.
До последнего времени предполагалось, что каталитическими свойствами обладали только полипептиды, в то время как нуклеиновые кислоты являлись только хранителями генетической информации. Однако недавно было показано, что нуклеиновые кислоты, в частности РНК, сами по себе обладают каталитическими свойствами. Были обнаружены две группы митохондриальной РНК грибов и группа ядерной РНК простейших, у которых сплайсинг обеспечивается самими РНК (self-splicing). Это отражает три возможных эволюционных пути автокаталитических реакций. Сплайсинг интронов группы 1 происходит в отсутствие белка и источников энергии, но требует присутствия гуанозина. При сплайсинге ядерной РНК образуется не кольцо, а лариат. По-видимому, соответствую щиеинтроны содержат каталитические центры или же эту функцию осуществляют так называемые ядерные РНК (И1 и И2). Во всяком случае эти свойства РНК иго-новому освещают происхождение жизни.
Для дальнейшего их перехода к первичным живым формам было необходимо появление матричного синтеза полипептидных цепей из имевшихся в «первичном бульоне» свободных аминокислот, а такой синтез стал возможным лишь с появлением транспортных РНК (тРНК). Каждый вид тРНК обладает двойной специфичностью — способностью узнавать определенный кодон в нуклеиновой кислоте и способностью связываться с определенной аминокислотой. Вместе с тем разные тРНК имеют общие свойства: все они состоят примерно из 80 оснований, имеют одинаковую 3'-концевую последовательность ССА, сходную конфигурацию и другие свойства, обеспечивающие возможность образования между аминокислотными остатками пептидных связей. Появление тРНК означало и появление генетического кода, так как каждому кодону полинуклеотидной цепи теперь соответствовал определенный антикодон тРНК, а каждый вид последней связывался с определенной аминокислотой.
На ранних стадиях химической эволюции репликация нуклеиновых кислот и синтез полипептидов происходили, вероятно, без органического катализа, хотя уже на этом этапе, по- видимому, существовали источники энергии типа АТФ. Однако появление полипептидов с ферментативными свойствами -резко ускоряло матричный синтез полинуклеотидов и белков и знаменовало собой переход от химической эволюции к биологической [Fox S., Nakashjma Т., 1980]. К этому времени, по-видимому, относится появление белково-липидных мембран, отделявших первичные формы жизни от окружающей среды. Возможно, на этой стадии химической эволюции аутокаталитические процессы стали пространственно локализоваться на первичных мембранных белково-липидных структурах с участием полисахаридов и полинуклеотидов. Моделью таких структур являются коацерваты, в которых могли протекать гетерокаталитические реакции [Опарин А. И., Гладилин К. Л., 1980]. Здесь кончается химическая эволюция и начинается эволюция биологическая (рис. 1).
В результате матричного воспроизведения соответствующих молекул в определенных условиях происходит следующее: конкуренция размножающихся молекул за вещество, энергию, пространство; вариабельность матричных молекул; воспроизведение в матричных копиях всех вариантов; естественный отбор, т. е. сохранение и размножение лишь наиболее приспособленных. Иными словами, уже на этом уровне начинают действовать законы наследственности, изменчивости и эволюции органического мира, что было хорошо иллюстрировано экспериментами, проведенными Ч. Дарвиным в пробирке.
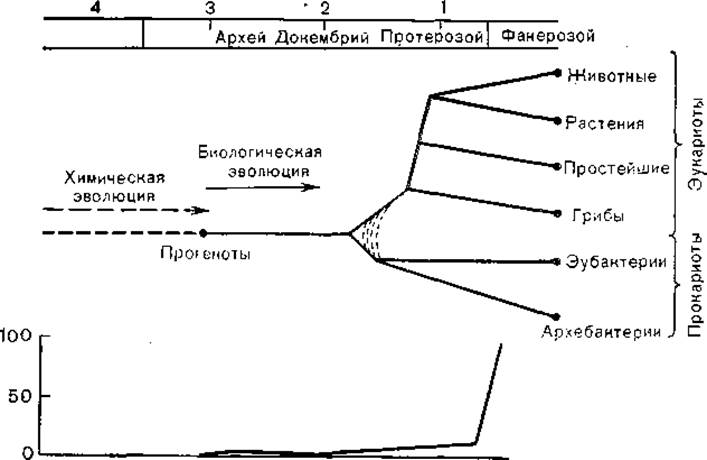
Рис. 1. Химическая и биологическая эволюция. По оси абсцисс — возраст (млрд лет); по оси ординат: вверху — эволюция органического мира, внизу — содержание кислорода в атмосфере (% к современному уровню).
Для эволюции биологических макромолекул характерна самоорганизация, которая уже имеет место на уровне примитивных нуклеиновых кислот. Рассматривая вопрос, что раньше возникло, белок или нуклеиновая кислота, М. Eigen (1971) считает его некорректным, так как это все равно, что спрашивать, что возникло раньше — информация или функция. Развивая концепцию самоорганизации материи, автор выдвигает общий принцип отбора и эволюции на молекулярном уровне, основанный на критерии устойчивости стационарных состояний по нелинейной термодинамической теории. Эволюция представляется неизбежным событием, если задано присутствие определенного вещества с определенными автокаталитическими свойствами и если поддерживается такая величина порога свободной энергии, которая необходима для компенсирования стационарного производства энтропии. Эта теория дает количественную основу для постановки лабораторных экспериментов по эволюции и правила построения простых молекулярных моделей, соответствующих возможным предшественникам живых клеток.
Для биологических полимеров характерна способность к самосборке. Этими свойствами, по-видимому, уже обладали предшественники биологических полимеров, образовавшихся в «первичном бульоне», в частности, протеноиды, которые легко формируют сферические (микросферы) и другие образования, характеризующиеся относительной стабильностью и определенной упорядоченностью внутренней структуры. Граничные их структуры напоминают клеточные мембраны. Вероятно, подобного рода самособранные образования и дали начало протоклетке (схема 2).
В статьях А. П. Руденко (1969, 1970) и Л. Б. Меклера (1980) на пути образования протоклетки отмечены следующие процессы или стадии: возникновение упорядоченности в белках в отсутствие макромолекул и современного генетического кода; возникновение ферментов, участвующих в их синтезе; возникновение обмена веществ в отсутствие клеток, «наделенных» обменом; возникновение клеток в отсутствие других клеток, которые могли дать им начало; возникновение систем аккумулирования энергии; возникновение генетического кода.
Предполагают, что первичные формы жизни содержали в качестве генетического материала РНК. ДНК появилась позже и с ее появления произошло разделение функций между ДНК и РНК в том виде, в котором они существуют в настоящее время: ДНК является хранителем генетической информации, а из двух видов РНК мРНК кодирует генетическую информацию с ДНК, а тРНК переводит генетический код с языка нуклеинового на язык белковый.
Появление рибосомной РНК (рРНК) и возникновение рибосом целиком относятся к ранним этапам биологической эволюции, и все сохранившиеся до нашего времени древнейшие формы жизни обладают рибосомными системами синтеза белков. Стабильность структуры тРНК и рРНК на протяжении нескольких миллиардов лет эволюции свидетельствует об исключительно стабилизирующем характере естественного отбора в этом случае.
Предполагают, что переход от протоклетки (proto-cell) к прокариотной клетке произошел в. промежутке 3—4 млрд лет назад. Предшественниками ископаемых бактерий были прогеноты. Ранние прокариотические клетки существовали в восстановительной атмосфере и нынешние метаногенетические бактерии, по-видимому, являются прямыми их потомками. Первоначальные прокариоты обладали способностью к анаэробному брожению. Затем появилась примитивная форма бактериального фотосинтеза, затем более сложная форма фотосинтеза типа растений (сине-зеленых водорослей) и лишь впоследствии аэробное дыхание.
Образования, проходившие долгий путь от неживой к живой материи, обозначаются как эобионты, более близкие предшественники ископаемых бактерий (более 3 млрд лет назад), как протоклетки, или прогеноты. По данным S. Fox соавт. (1977), изучавших гомологии 185 рРНК в целях исследования филогенеза прокариотов, первоначальные прокариоты дали начало трем эволюционным линиям: уркариотам (Urkaryotes), ставшим симбионтами, потомками которых являются цитоплазматические компоненты эукариотов; архебактерии (Archaebacteria), потомками которых являются современные метанобактерии, и эубактерии (Eubacteria) —предки современных бактерий.
СХЕМА 2. ЭВОЛЮЦИЯ САМОСБОРКИ
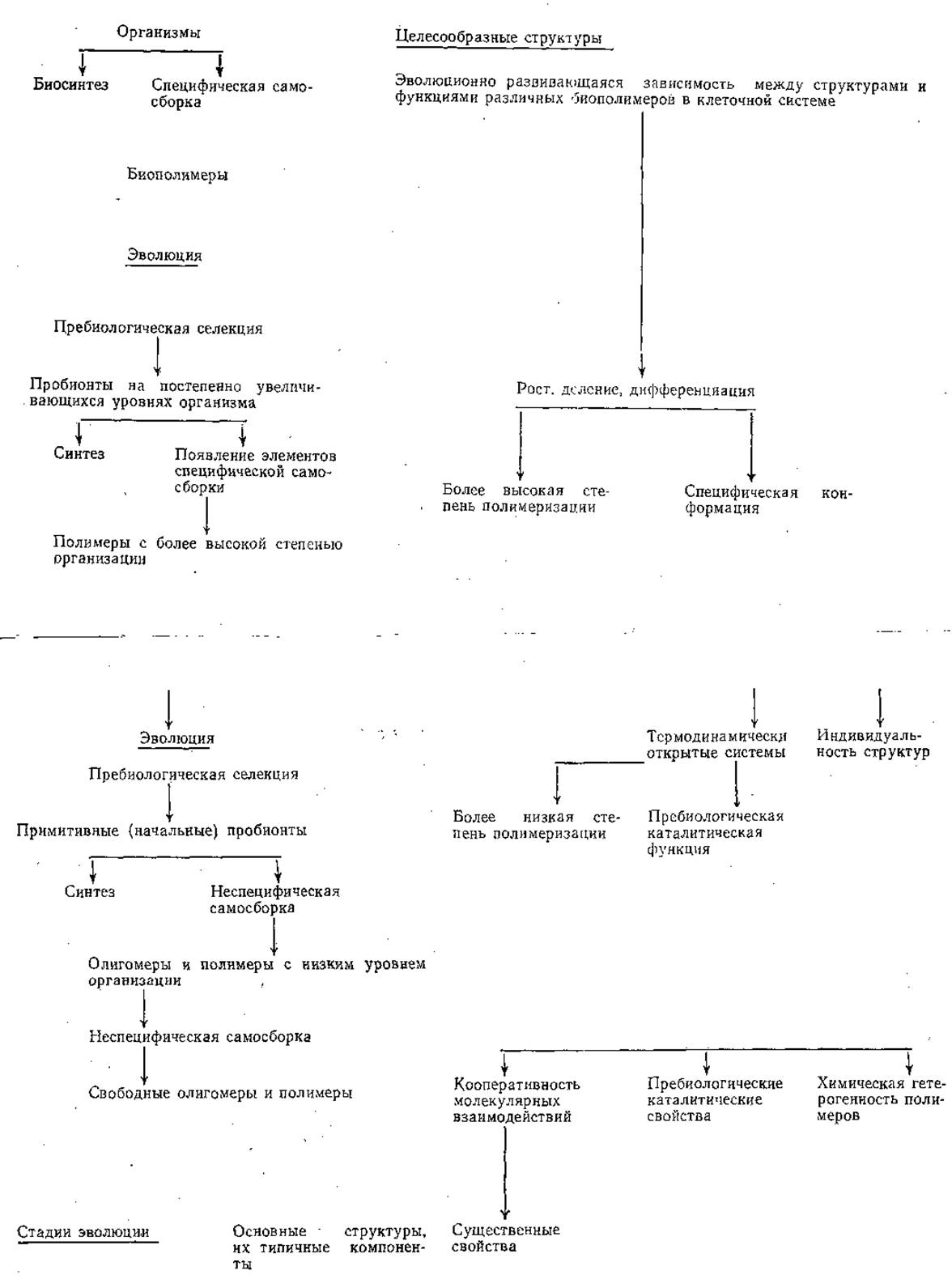
В процессе биогенеза, или биопоэза, различают стадии коацервации, формирования мембран, появления метаболизма, самовоспроизведения (редупликация). Уже на данных стадиях могли появиться мутации, приобретавшие новую способность — способность к органическому фотосинтезу. Этот новый образ жизни давал большие преимущества по сравнению с анаэробиозом. Глобальным результатом органического фотосинтеза стало накопление в атмосфере свободного кислорода, которое продолжалось около 2 млрд лет.
Смена восстановительной атмосферы на кислородную началась между средним и поздним докембрием, около 1,8 млрд. лет назад, а кислородная атмосфера сформировалась 1,4 млрд лет назад. Таким образом, если жизнь появилась на Земле 2,7—3,2 млрд лет назад, то потребовалось 0,9—1,4 млрд лет, чтобы появились организмы, использующие фотосинтез.
Нарастание содержания кислорода в атмосфере до 1% современного уровня повлекло за собой многие последствия, поскольку уже при такой его кондентрации поглощалась самая опасная для жизни часть ультрафиолетового излучения. Предполагают, что такая атмосфера образовалась в начале кембрия и этот период соответствует «взрыву жизни», нашедшему отражение в изобилии ископаемых. Если до этого обитаемым был только слой воды не глубже 50 м (куда еще проникает солнечный свет) и не мельче 10 м (куда проникало смертельное жесткое ультрафиолетовое излучение), то с изменением состава атмосферы за счет увеличения в ней кислорода для жизни стали доступны и поверхностные слои водоемов, а впоследствии стало возможным и завоевание суши. Одновременно с нарастанием количества кислорода в атмосфере уменьшилось количество двуокиси углерода, поступавшей из недр земли и являвшейся основным источником образования кислорода.
Предполагаемая история атмосферного кислорода и двуокиси углерода освещена в работе W. Rutten (1971). По его данным, содержание кислорода в атмосфере, составлявшее 0,001% от современного уровня, начало повышаться около 3,2 млрд лет назад, достигнув к 2,7 млрд лет назад 0,01% от современного, 400 млн лет назад 0,1% от современного и 200 млн лет назад современного уровня. Одновременно с этим уменьшалось содержание двуокиси углерода, которое в докембрии было 10-кратным по сравнению с современным. При этом в периодах интенсивных горообразований содержание двуокиси углерода резко возрастало вследствие выбросов его из земных недр в атмосферу.
Первые следы жизни — микроскопические биогенные отложения и молекулярные ископаемые — обнаруживаются в раннем и среднем докембрии, возраст их определяется более 2,7 и даже 3,2—3,7 млрд лет. На репликах со шлифов в электронном микроскопе обнаружены глобулярные и нитчатые образования, напоминающие клетки бактерий и сине-зеленых водорослей. Эти организмы обладали метаболизмом, характеризовавшимся секретированием карбоната кальция (строматолиты). Если считать возраст Земли более 4,5 млрд лет, то первые кокковые и нитчатые бактерии обнаружены в осадочных породах 3,5 млрд лет назад, а отделение эукариотов от прокариотов произошло 2—3 млрд лет назад. Гипотетический общий предок имел геном, составляющий 74 генома современных прокариотов. В позднем докембрии уже регулярно обнаруживаются микроорганизмы, напоминающие бактерии и сине-зеленые водоросли. В раннем кембрии уже имеются многочисленные ископаемые, относящиеся к многоклеточным организмам.
Итак, химическая эволюция продолжалась более 1 млрд лет и завершилась в среднем докембрии (позднем архее) образованием первых клеточных организмов — предшественников прокариотов.
Что же касается эукариотов, то в настоящее время почти общепризнано, что они, произошли от симбиоза двух прокариотических форм, из которых большая клетка была гетеротрофным анаэробам, а меньшая — прокариотом, способным к дыханию. В дальнейшем этот симбиоз стал обязательным, причем в него могли включиться несколько разных прокариотов. Клетка-хозяин постепенно «отделила» ядро, а потомками симбионтов являются митохондрии, хлоропласти и, возможно, центриоли с аппаратом митотического веретена [Sagan L., Margulis L., 1967]. Структуры типа эукариотов впервые обнаруживаются в отложениях давностью 1,5 млрд лет назад.
Серьезным доводом в пользу симбиотического происхождения эукариотов является сходство рибосомного аппарата митохондрий и хлоропластов эукариотов с таковым прокариотов, в частности, большая гомология 16S РНК митохондрий и хлоропластов с 16S РНК бактерий и отсутствие такой гомологии с цитоплазматической РНК с молекулярной массой 18 000 [Bonen L. et al., 1977]. Рибосомы митохондрий и хлоропластов чувствительны к хлорамфениколу, как рибосомы прокариотов, в то время как цитоплазматические рибосомы эукариотов нечувствительны к этому антибиотику. Эта точка зрения разделяется, однако не всеми. В частности, в настоящее время накоплено много фактов, указывающих «а репродукцию ряда вирусов в хлоропластах и митохондриях. Основываясь на этих данных, выдвинуто предположение о существовании вирусов, использующих ферментные системы этих органелл («митофаги», «хлорофаги»), которые реплицировались в эволюционных предшественниках пластид и митохондрий. При этом возможны две модели эволюционного процесса: прокариотная ДНК сохранилась автономной в органеллах, происшедших от соответствующих прокариотов — симбионтов эукариотических организмов, или ДНК эндоплазматических симбионтов вошла в состав ядерной ДНК эукариотов. По-видимому, обе модели были реализованы в ходе эволюции эукариотов, причем в случае транспорта прокариотных генов в ядро эукариотов векторами могли быть упомянутые выше гипотетические вирусы.
Имеются, однако, и критические замечания по концепции об экзогенном эндосимбиотическом происхождении органелл эукариотической клетки [Серавин Л. И., 1986]. Ей противопоставляется концепция об аутогенном (эндогенном) их происхождении путем эволюции поверхностной мембраны, цитоплазмы и нуклеоида. К сожалению, автор, выявляя трудности объяснения некоторых фактов (строение ДНК, тРНК митохондрий и хлоропластов), обходит очень важный вопрос о гомологии нуклеиновых кислот органелл и прокариотов, поэтому его доводы нельзя признать убедительными. К томy же концепция об эндогенном происхождении органелл не пытается даже объяснить факты, не укладывающиеся в ее рамки.
Предполагают [Woese С., Fox G., 1977], что прокариоты и эукариоты имеют общего предка — прогенота (progenote). Если считать, что жизнь на Земле началась более 3 млрд лет назад, то реконструкция эволюции на основании анализа ископаемых возможна лишь для последних 500 млн лет. В течение 1—2 млрд лет продолжался «век прокариотов», хотя и в это время уже могли существовать примитивные эукариоты. «Расцвет» их начался, вероятно, позже того как установился симбиоз их с прокариотами, приведший к появлению современных форм эукариотов. Вероятно, Polomyxopalistris является прообразом примитивных эукариотов. Этот организм не имеет митохондрий и митотического аппарата, не проявляет 9+2-фибриллярных структур, не имеет мембран аппарата Гольджи, однако содержит прокариотический симбионт. Очевидно, Eukaryotae включают в себя органеллы прокариотического происхождения. С другой стороны, метаногенные бактерии настолько отличаются от остальных бактерий, что должны быть выделены в отдельное царство. В этом случае Procaryotae состоят из двух давно разошедших ветвей — архебактерий и эубактерий. Метаногены должны быть отнесены к архебактериям.
С. Woese (1979) оспаривает предположение о том, что митохондрии произошли 1—2 млрд лет назад от симбионтов — аэробных бактерий. Он считает, что эндосимбиоз имел место значительно раньше, в анаэробном периоде, и привел к образованию фотосинтетической органеллы, аналогичной-современному хлоропласту.
Обобщая длительную дискуссию по этому вопросу, М. Gray и W. Doolittle (1982) приходят к выводу о самостоятельной и длительной, эволюции двух царств эукариотов—Archaebacteria и Eubacteria. Потомками первой ветви явились Hatobacteria, Methanogenes, Thermoplasma, Sulfvibrio и др., а потомками второй ветви — все остальные бактерии, включая Cyanobacteria, а также эндосимбионты эукариотов, ставшие впоследствии их органеллами, — пластиды (хлоропласты), митохондрии. Авторы приводят много доказательств этому на разных уровнях: гомология рРНК (16S, 5S) органелл и бактерий, цитохрома Ce(t), ферродоксина, рибосомных белков, тРНК, инициация с формилметионинового кодона, сходство факторов; трансляции, факторов ppGpp и ppGppp и др. Но в то время как пластиды мономорфны и есть основания -предполагать, что все они имеют монофилетическое происхождение и возникли от предков нынешних сине-зеленых бактерий, митохондрии грибов, простейших, растений и животных сильно различаются между собой и следует скорее думать о полифилетическом их происхождении, притом более древнем, нежели у хлоропластов. М. Gray и М. Doolittle выделяют главные положения, по которым (которыми) эукариоты отличаются от прокариотов.
1. В то время как трансляция и транскрипция у прокариотов «спарены», у эукариотов они разобщены и генетический аппарат размещен в множественных линейных хромосомах с множественными началами репликации. 2. Деятельность ДНК регулируется гистонами, которых нет у прокариотов. 3. Вместо оперонов, как у бактерий, у эукариотов существует система промоторов и терминаторов, ограничивающих синтез полицистронной мРНК. 4. Имеется три специфические РНК-полимеразы для генов больших рРНК, для генов, кодирующих белки, и для низкомолекулярных РНК (5S рРНК, тРНК). 5. Имеются система экзонов, интронов и сплайсинг. 6. Имеются гены 25S, 18S и 5S РНК. 7. мРНК имеет кэп-структуры на 5'-конце и поли (А) на 3'-конце. 8. В ДНК имеются множественные повторы, лишенные кодирующего смысла.
Авторы полагают, что геном эукариотов возник 1—2 млрд, лет назад из древнего Эубактериального генома. Он претерпел быструю и фундаментальную структурную реорганизацию. Возможными предками современных органелл стали цианобактерии, микоплазмы, микобактерии, актиномицеты и родобактерии. Возможно, что именно архебактерии явились предками нынешних эукариотов, в частности Thermoplasma или TIalobacteria, поскольку по ряду параметров они более сходны с эукариотами, чем эубактериями [Fox S. et al., 1980].
Одним из возможных путей обогащения генома эукариотов является образование псевдогенов, которое было детально изучено у глобиновых генов мышиных клеток. Эти гены прерывисты и имеют два интрона разной величины, которые при сплайсинге мРНК удаляются. В этих же мышиных клетках обнаружены псевдогены — копии экзогенов истинных генов, у которых отоутствуют нитроны. Кроме того, у них наблюдаются небольшие делеции, в результате которых псевдогены не функционируют в качестве генов [Vanin Е. et al., 1980]. Псевдогены детергированы в трех разных хромосомах [Leder А. et al., 1981]. Предполагают, что псевдогены возникли путем обратной транскрипции мРНК, у которой интроны были удалены в результате сплайсинга, и псевдогены каким-то образом контролируют работу нормальных генов. В пользу появления их в результате обратной транскрипции мРНК свидетельствует наличие в псевдогенах поли (А) последовательностей [Hollis G. et al., 1982]. Наличие псевдогенов может являться источником мутаций, приводящих к возникновению новых генов.
Псевдогены эволюционируют необычайно быстро, что было показано при изучении псевдогенов глобина и 2/2 цепей иммуноглобулина [Miyagata Т., Hayashida Н., 1981].
Для построения филогенетических древ эволюции организмов в прошлом и настоящем широко применяется метод сравнительной морфологии, при котором параллельно изучают как ныне оуществующие виды, так и вымершие, ископаемые, виды. Естественно, чем далее мы продвигаемся в глубь истории, тем более бедными являются ископаемые и тем менее сохраняются морфологические особенности вымерших организмов. С развитием молекулярной биологии стало возможным восстанавливать филогенетические древа на основании изучения гомологии белков и нуклеиновых кислот. Именно таким образом было показано, что сине-зеленые водоросли ближе к бактериям, чем к другим водорослям, метанобактерии дивергировали от общего предка с бактериями и сине-зелеными водорослями и цитоплазматические компоненты эукариотов (митохондрии, хлоропласты) являются дериватами прокариотов, ставших симбионтами древних эукариотов.
Общий ход эволюции органического мира заключается в возрастании биологического совершенства организмов. Согласно А. Н. Севердову (1939), существуют два основных пути эволюционного процесса: ароморфозы и идиоадаптации. При ароморфозах биологическое совершенство количественно и качественно поднимается на более высшую ступень, плоскость. Примерами могут служить последовательные появления первичных хордовых, рыб, земноводных, пресмыкающихся и птиц, млекопитающих. При идиоадаптации появляются признаки, которые могут случайно оказаться носителями полезных свойств и влекут за собой перестройку (морфологическую и физиологическую) разных органов.
Например, у предков высших позвоночных амниот (классы пресмыкающихся, птиц и млекопитающих) развивался эффективный механизм вентиляции легких посредством работы грудной клетки. Этот ароморфоз должен рассматриваться именно- как ключевой, поскольку его появление повлекло за собой длинную цепь важных перестроек организации (и как следствие образа жизни), которую можно схематически представить следующим образом. Исчезла необходимость кожного- дыхания (последнее при недостаточной эффективности вентиляции легких у земноводных было важным дополнением легочного); в покровах усилились процессы ороговения эпидермиса, что защитило организм от обезвоживания и обеспечило лучшую механическую защиту; одновременно открылись возможности для последующей сложной дифференциации производных покровов (развитие роговых чешуй рептилий, перьев птиц, волос млекопитающих); исчезла необходимость смешивать в желудочке сердца кровь от легких и от кожи (у земноводных это было необходимо опять-таки в связи с дыхательной ролью кожи) й в результате появилась возможность для разделения артериальной н венозной крови в сердце внутрижелудочковой перегородкой, что в свою очередь привело к перестройкам отходящих от сердца артериальных стволов и повысило уровень метаболизма и развития гомойотермии; утрата ротоглоточной полостью и подъязычным аппаратом роли дыхательного насоса позволила изменить общую конфигурацию черепа. Все эти изменения, сами по себе значительные и важные, стали возможными лишь после осуществления ключевого ароморфоза.
Другие авторы связывают появление ароморфозов с резкими изменениями внешней среды, активно селектирующими мутации, которые в стационарных условиях устраняются, так как в этом случае имеет место стабилизирующий отбор [Шмальгаузен И. И.,- 1969]. При освоении группой организмов новой адаптивной зоны имеется некоторая промежуточная стадия, когда уже утрачена приспособленность к прежней зоне, но еще не развилась приспособленность к новой. Эта неадаптивная фаза быстро преодолевается под влиянием естественного отбора, особенно если данная группа организмов имеет необходимые предадаптации, на основе которых развиваются приспособления к новой адаптивной зоне. Такая эволюция обладает как бы скачкообразным характером (квантовая эволюция), и темпы ее во много раз превышают обычные для данного таксона темпы эволюции.
Такие условия создаются при изоляции небольшой части популяции, когда на генофонд данной группы действует не стабилизирующий отбор, а отбор полезных для данной группы мутаций. В этом случае перестройка генофонда происходит настолько быстро, что ее обозначают как генетическую революцию [Mayr F., 1963].
L. Mettler и Т. Gregg (1972) следующим образом формулируют применение законов Менделя к популяционной генетике. В отсутствие сил, вызывающих изменение «генных основ», большая популяция со случайным скрещиванием будет находиться в состоянии генетического равновесия. Эволюция представляет собой постепенное изменение генных признаков, за счет таких процессов, как отбор, мутации, миграция и дрейф генов. При постоянстве условий среды эти силы стремятся привести популяцию в состояние генного равновесия, при изменении условий среды популяция, стремится к новому равновесию.
Отбор внутри популяции может быть стабилизирующим, направленным и дизруптирующим. Если первый тип отбора удерживает популяции вблизи средних значений ее количественных признаков, а второй тип приводит к сдвигам этих средних значений, то третий тип отбора обеспечивает дивергенцию субпопуляций и в конечном счете возникновение новых видов. Дрейф генов, т. е. случайные колебания частот генов в небольших изолированных популяциях, сам по себе не играет большой роли в эволюции, пока данная популяция остается малочисленной или изолированной. Такие популяции нередко вымирают, если условия среды изменяются в неблагоприятную -сторону. Если такие популяции сливаются с другими, свободный обмен генами быстро уравновешивает дифференциацию, развивающуюся в колонии за время ее изоляции. Таким образом, поток генов и его последствия не способствуют эволюции [Mayr F., 1963]. Однако, если изменения условий среды приводят к расширению экологической ниши данной популяции, возникают огромные потенциальные возможности для ее эволюции.
Как указывают L. Mettler и Т. Gregg (1972), для того чтобы существовать, популяция должна быть приспособлена к среде ее обитания. Это значит, что большая часть ее особей должна иметь такие генотипы и фенотипы, которые обеспечивают ей существование в данных условиях. А для дальнейшего существования популяция должна сохранять свою приспособленность к среде, изменяя свою генетическую структуру при неизбежных изменениях среды. Иными словами, в популяции должны возникать и воспроизводиться новые генотипы и фенотипы, приспособленные к изменившимся условиям, а это возможно при постоянно происходящей генетической изменчивости популяции. В пределах этой изменчивости определенное место занимает изменчивость, «поставляющая» плохо приспособленные фенотипы, которая обозначается как генетический груз популяции [Haldane J., 1957]. Последний включает в себя мутационный груз, сбалансированный груз и субституционный груз.
Мутационный груз — это доля общего генетического груза, которая возникает за счет мутантных аллелей. Естественный отбор обычно направлен против этих аллелей, одна ко в определенных условиях может им способствовать. Так, например, в неблагополучных по малярии районах Африки распространена серповидно-клеточная анемия — результат мутационной замены одной аминокислоты в молекуле гемоглобина. Лица с такой заменой устойчивы к малярии, и отбор данного признака доминирует, а платой за него является болезнь крови. При сбалансированном грузе характерно существование множественных аллелей, которые выгодны в разных условиях, у особей разного пола, на различных стадиях развития и т. п. Примером могут служить существование многочисленных изоферментов, а также сохранение полиморфизма благодаря сверхдоминированию. В ряде случаев в связи с изменениями условий среды «неблагоприятные» и «благоприятные» аллели меняются своими местами и отбор идет направленно, подавляя прежде «благоприятный» и поэтому распространенный аллель и стимулируя прежде «неблагоприятный» и подвергавшийся в результате этого вытеснению аллель. В определенные периоды своего развития популяция становится плохо приспособленной и возникает груз, называемый переходным, или субституционным.
По расчетам J. Haldane (1957), для замещения одного аллеля необходимо в среднем 300 поколений и в 30 раз больше смертей, чем имеется в каждом поколении. Если предположить, что для образования нового вида достаточно 1000 замещений, то для этого потребуется 30 000 поколений. Такая расчетная скорость эволюции в общем соответствует скорости для многих видов млекопитающих на основании палеонтологических данных. Однако применительно к эволюции человека эти расчеты оказались непригодными. Если считать, что за 1 млн лет эволюции человека сменилось 50000 поколений (среднее время между поколениями 20 лет), то их хватило бы на замещение 166 генов. Между тем различия между человеком и его обезьянеподобными предками должны определяться гораздо большим числом генов. Таким образом, скорость эволюции человека была в 6—7 раз больше расчетной. Эти выводы вытекают из анализа скорости эволюции митохондриальной ДНК у приматов [Brown F. et al., 1979]. При оценке скорости эволюции необходимо иметь в виду, что предпосылкой видообразования являются изоляция ограниченной части популяции и интенсивный инбридинг. Подобного рода обстоятельства создаются при резких изменениях условий внешней среды, приводящих к вымиранию части популяции. В качестве примера можно привести эволюцию человека, которая шла интенсивно на протяжении последнего миллиона лет плейстоцена, когда было четыре периода оледенения. За это время появились и исчезли «предшественники» человека питекантропы и неандертальский человек и сформировался современный человек. При нынешней численности огромная популяция человека гетерогенна и именно поэтому не имеет шансов на дальнейшую эволюцию.
Считают, что для морфологической дифференциации подвидового порядка требуется не менее 300 поколений, однако скорость эволюции у разных животных неодинакова. Так, некоторые млекопитающие, проникшие из Северной Америки в Южную в плиоцене и плейстоцене, т. е. 1—2 млн лет назад, образовали эндемические роды, но ни одни из них не дали эндемического семейства. Средняя продолжительность существования рода у пластинчатожаберных (Pelecypoda) равна 78 млн лет, тогда как у хищников (Carnivora) она достигает лишь 6,5 млн лет. К наиболее, существенным факторам, которые могут влиять на скорость и пути эволюции, относятся изменчивость, частота мутаций и их характер, скорость смены поколений, размеры популяций и естественный отбор [Simpson G., 1945, 1953]. Однако, как указывает G. Simpson, эволяция на основе существующей изменчивости — самоограничивающийся процесс, который не может выйти за пределы, примерно соответствующие уровню видообразования.
Одно из направлений современных эволюционных концепций представлено нейтралистской теорией М. Kjmura (1968), связанной с развитием молекулярной биологии [Волькенштейн М. В., 1983]. Известно, что подавляющая часть мутаций нуклеиновых кислот либо не ведет к замене соответствующей аминокислоты (если произошла замена 3-го нуклеотида в кодоне), либо ведет к замене на сходную аминокислоту (иначе мутация будет летальной). В этом заключается высокая помехоустойчивость генетического кода. В результате сохраняются активный центр молекулы белка и основа ее каркаса, в то время как аминокислотный состав белка может существенно меняться. Поэтому белки с разной первичной структурой молекулы могут иметь сходное пространственное строение, и это связано с тем, что большинство мутаций являются нейтральными.
В дальнейших работах М. В. Волькенштейна (1986) отмечалось, что, сохраняя основные положения теории эволюции
4. Дарвина, можно пересмотреть некоторые ее положения в свете молекулярной биологии, синергетики и теории информации. Автор подчеркивает, что видообразование могло идти не постепенно, а скачками (пунктуализм), и в этом случае ароморфозы и дезадаптации, по А. Н. Северцову, являются чередованиями пунктуализма и градуализма. Он подчеркивает обусловленность мутаций сложившимся строением организма, в связи с чем имеется определенная направленность эволюции («внутренняя канализация»), а многие мутации имеют неадаптационный характер. Идея направленности эволюции коррелирует с концепцией Н. И. Вавилова о гомологических рядах изменчивости.
С этим связаны дополнительные соображения о направленной эволюции, которая определяется уже сложившейся структурой организма, ограничивающей многие виды случайных мутаций, — соображения, высказанные Н. И. Вавиловым в рамках указанной концепции. Отсюда вытекает вывод, что многие признаки не имеют адаптированного значения, о чем указывал еще С. С. Четвериков.
Нейтралистская теория М. Кимуры сформулирована с учетом следующих принципов. 1. Для каждого белка скорость эволюции (число замещений аминокислот на один остаток в год) примерно одинакова, пока структура и функции данного белка существенно не меняются. 2. Функционально важные молекулы или их участки эволюционируют медленнее, чем функционально маловажные. 3. Менее опасные замены аминокислот происходят чаще, чем более опасные. 4. Дупликация генов всегда предшествует появлению гена с новой функцией. 5. Элиминация вредных мутаций и случайных фиксаций нейтральных мутаций происходят чаще, нежели отбор полезных мутаций.
Возможные причины гомологии аминокислотных последовательностей рассматриваются как результат либо эволюционной общности, либо молекулярной конвергенции. При этом подчеркивается, что большинство белков, особенно ферментов, возникло из ограниченного числа полипептидов.
Мысль о возможности молекулярной конвергенции вытекает также из сравнения бактериальных и митохондриальных цитохромов (с2, с). Сограоно Т. Meyer и соавт. (1986), когда структура и функции двух гомологичных участков белков достигают определенного соответствия, конвергирующие мутации становятся столь же частыми, как и дивергирующие.
Нам хотелось бы дополнить эти данные своими соображениями, которые мы формулируем как концепцию молекулярной конвергенции. Благодаря развитию молекулярной биологии выявилось существование химически сходных структур (нередко с разными функциями) в различных группах организмов, которые не имеют между собой даже отдаленных эволюционных связей. Наиболее ярким примером служит существование родопсина у бактерий и в зрительных органах низших и высших животных. Обычное объяснение эволюционной преемственности, например у тРНК, эволюция которых шла исключительно медленными темпами, здесь, естественно, отпадает. Отпадает и возможность переноса соответствующих, генов векторами-типа вирусов или плазмид, как это могло происходить с бактериальными токсинами. Приходится признать поэтому, что родопсин 'бактерий и родопсины зрительных клеток низших (осьминогов) и высших (позвоночных) животных были трижды независимо «изобретены».
В связи с соображениями о молекулярной конвергенции необходимо, по-видимому, рассмотреть данные о молекулярной мимикрии. При исследовании более 600 моноклональных антител, направленных против 11 разных вирусов, с белками (экстрактами) из 14 различных органов незараженных мышей, было показано, что 21 из них (3,5%) реагировали с белками, этих органов. Здесь же не может быть ни эволюционной связи, ни захвата клеточных генов, так как реакции эти были получены с разными вирусами (Коксаки, японского энцефалита, лимфоцитарного хориоменингита, кори, бешенства, везикулярного стоматита, герпеса, оспы). Отрицательные результаты были получены с вирусами денге и цитомегаловирусами человека и мышей. U. Jahnke и соавт. (1985) обнаружили декапептиды, гомологичные человеческому миелину, в белках вирусов кори, Эпштейна — Барр, гриппа А и В. Авторы полагают, что имеется связь между этой находкой и аллергическими энцефаломиелатами. Вероятно, во всех этих случаях выявлялись кластеры аминокислотных последовательностей, общие для вирусов и мышиных клеток, поскольку речь шла о своеобразной молекулярной конвергенции.
При толковании мутационных процессов (имеются в виду прежде всего точечные мутации) обычно подчеркивается не только их случайность (стохастический характер), но и почти бесконечное число возможных мутаций. На самом деле это далеко не так, потому что число возможных замен строго ограничено. Возьмем, например, антигенную детерминанту гриппа, состоящую, скажем, из 10 аминокислотных остатков. Если бы каждую аминокислоту можно было бы заменить любой другой (а к этому ведут случайные мутации), то число антигенных вариантов достигло бы астрономической цифры — 1020. На самом деле их не более немногих сотен или даже десятков, так как большинство замен (например, триптофан вместо аланина) ведет не к модификации, а к разрушению антигенной детерминанты, а иногда и летальной модификации всей молекулы гемагглютинина. Именно поэтому такая «простая» эволюционная находка, как АТФ, сохранилась на всех стадиях эволюции органической жизни на Земле как универсальный аккумулятор энергии, а ригидная структура полифункциональных и полидоменных тРНК претерпела незначительные изменения за миллиарды лет эволюции. Именно поэтому же трижды независимо «изобретенный» родопсин оказался сходным по структуре у столь разных организмов, как бактерии, осьминоги и позвоночные. Образование этих структур было не только детерминированным, но и неизбежным, так как только они могли выполнить функции, вызванные к жизни эволюционным процессом.
Своеобразную концепцию эволюции генов развивает R. Dawkins (1976). Он называет свою концепцию фундаментальным законом эгоизма генов (gene selfishness). Автор считает ошибочными взгляды, будто живые творения «evolve to do things for the good of species», поскольку эти представления едва ли соответствуют дарвиновской теории, согласно которой индивидуальные члены вида в первую очередь соблюдают их собственные «selfish interests». По его предположению, на ранних стадиях эволюции живой материи, на стадии химической эволюции в результате случайных процессов появился репликатор — молекула ДНК, делающая свои копии. В результате мутаций (ошибок) первобытный суп (the primeral soup) был наполнен неидентичными репликами, что сопровождалось эволюцией их в сторону удлинения (toward greater longerity), а также увеличения скорости репликации и стабилизации. С этого времени начали действовать «соревнование» и естественный отбор. В результате возникли современные гены и их ансамбли с механизмами их репликации, который автор обозначает как «the gene machine», а многоклеточные организмы как «колонии генов». Система генов обладает стабильностью и относительной автономией. В итоге эволюция органического мира является прежде всего эволюцией генов.
Неотвратимость прогрессивной эволюции можно видеть на примере появления разума, который является далеко не случайным феноменом. Образно выражаясь, природа трижды пыталась создать разум и это ей удалось только после третьей попытки. Первой «попыткой» явилось развитие нервной системы у осьминогов и кальмаров, которых недаром называют приматами моря. Разум не мог развиться — мешали зависимость от водной среды, отсутствие орудий и другие факторы, несмотря на то что у этих животных уже появились условные рефлексы и индивидуальное, сходное с разумным, поведение, а нервная система получила значительное развитие. Второй «попыткой» были насекомые. Однако их малые размеры (что ограничивалось наружным скелетом), ограниченная возможность увеличения объема нервной системы обрекли на неудачу и эту «попытку», «венцом» которой стали колониальные насекомые — муравьи, пчелы, термиты, относящиеся, кстати сказать, к разным таксономическим группам. Поэтому в этом случае правильно говорить не об одной, а о трех неудачных «попытках» создать разум.
Появление разума стало возможным благодаря возникновению и эволюции позвоночных и их наиболее прогрессивной ветви — млекопитающих. У них было все необходимое: удвоенный по сравнению с пресмыкающимися геном с избытком генетического материала, внутренний скелет, значительная независимость от окружающей среды вследствие развития теплокровия. Поэтому появление приматов, а затем человека разумного было не только возможно, но и неотвратимо, в то время как другие ответвления (например, дельфины, слоны, хищники) в плане возможности появления разума остались тупиками вследствие специализации.
С развитием молекулярной биологии для исследования эволюционных связей между разными организмами стало широко применяться изучение гомологии белков и нуклеиновых кислот — аминокислотной последовательности полипептидных цепей и нуклеотидной последовательности РНК и ДНК. Кроме того, используют дополнительные приемы для учета скоростей эволюции. С помощью этих методов можно не только выявлять филогенетические связи между разными организмами, но и учитывать скорости эволюции при построении эволюционных древ.
Так, при сравнительном изучении гистонов Н4 простейших и высших эукариотов показано, что различия пептидных цепей этих гистонов менее выражены у животных и растений, нежели у простейших, с одной стороны, животных и растений — с другой. Это позволяет прийти к заключению, что дивергенция Protozoa и высших эукариотов произошла существенно ранее дивергенции растений и животных. На основе данных изучения секвенций, получаемых после обработки РНКазой рибосомной РНК, митохондрий, хлоропластов, бактерий и РНК рибосом цитоплазмы животных и растений, было показано, что РНК рибосом хлоропластов и митохондрий имеют большую степень гомологии между собой и РНК бактериальных рибосом и значительно меньшую — с РНК рибосом цитоплазмы растений и животных. Это позволяет сделать вывод о прокариотическом происхождении РНК митохондрий и хлоропластов [Bonen L., Doolittle D., 1979].
G. Fox и соавт. (1977) при сопоставлении структуры 16 рибосомальных РНК у прокариотов пришли к заключению, что метаногеиные бактерии филогенетически далеки от истинных бактерий; к ним ближе сине-зеленые водоросли, а позже произошло разделение истинных бактерий на бацилл и вибрионов. Основываясь на изучении рибосомальной РНК, О. Woese и G. Fox (1977) выделяют у прокариотов три первичные линии развития: первоначальные формы прокариотов — Urkaryotes, которые дали начало цитоплазматическим компонентам эукариотов (митохондриям и хлоропластам); Archaebacteria, потомками которых являются метаногенные бактерии; Eubacteria, давшие начало современным разнообразным группам бактерий. Н. Hori и S. Osava (1979) сравнили вторичную структуру 55 РНК 54 видов, относящихся к прокариотам и эукариотам, — от бактерий до человека. На основании полученных данных было сконструировано филогенетическое древо. Для этого, во-первых, была рассчитана степень замены нуклеотидов, во-вторых, положено, что интенсивность замены пропорциональна числу лет, прошедших со времени эволюционной дивергенции двух молекул от их общего предка. По этим расчетам время дивергенции про- и эукариотов составляет 1,8 х 109лет, а дивергенции человека и дрожжей—1,2 х 109 лет. Эти же расчеты показывают, что грибы отделились от животных и растений несколько ранее, чем разделились, животные и растения, хотя возможно, что сначала произошло отделение растений от грибов и животных, а затем разделение двух последних. Несколько неожиданным является положение Helobacterium cutirubrum: в то время как бактерии происходят из одного ствола, эти бактерии скорее происходят от ветви эукариотов.
Если исходить из дивергенции рибосомной 55 РНК, то филогенетическое древо выглядит следующим образом (рис. 2). Линии эволюции про- и эукариотов являются самостоятельными и ведут начало от гипотетического общего предка. В эволюции эукариотов наиболее ранними ответвлениями являются протисты (Euglena), вслед за чем последовало значительное повышение радиации, что обусловило последовательное выделение грибов, растений и животных. Изучение филогении цитохрома с позволило также восстановить филогенетическое древо этого белка, напоминающее эволюцию соответствующих таксономических групп организмов. За основу было взято минимальное число мутационных различий в этом белке. Грибы ранее всего дивергировали от общего предка с животными. Значительно позже произошла дивергенция беспозвоночных и позвоночных, затем отделились рыбы, пресмыкающиеся, затем произошла дивергенция млекопитающих я птиц и, наконец, человека и обезьян.
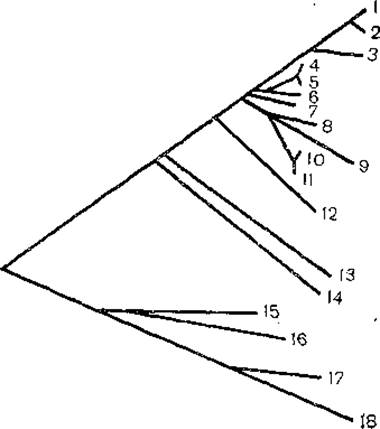
Рис. 2. Межцарственная филогения, распределенная по сходству последовательностей малой субъединицы рибосомной РНК.
1— крыса; 2 - Х. laevis; 3 — A. salina; 4 — 2. mays; 5 — рис; 6 — A. castelanii; 7 — S. cerevisiae; 8 — P. tetraurelia; 9 — T. thermophila; 10 — O. nova; 11 — S. pustulata; 12 — D. discoideum; 13 — T. brucei; 14 — E, gracilis; 15 — S. solfataricus; 16 — H. volcanii; 17 — A. nidulans; 18 — E. coli.
Открытие феномена сплайсинга [Berget S. et al., 1977] дало возможность по-новому оценить события в ранней эволюции, поскольку наличие нитронов и экзонов было установлено не только у низших, но и у высших эукариотов. Высказывалось предположение, что экзоны кодируют определенные до, мены белков, в то время как нитроны могут стать объектом быстрой эволюции. В соответствии с этими соображениями была предложена модель молекулярной эволюции [Darnell J., Doolittle W., 1986] (см. схему 1), причем на ранних стадиях развития отщепление нитронов и лигирование экзонов обеспечивались самой РНК.
Наиболее древние окаменелости структур, напоминающие сине-зеленые водоросли, обнаружены в породах, которые имеют возраст около 3,1 млрд лет. Структуры этого же типа встречаются и в более молодых породах с возрастом около 0,8 млрд лет. Вероятно, 1,5 млрд лет назад появились первые многоклеточные животные (губки). Таким образом, потребовалось 2 млрд лет, чтобы эволюция первоначальных биологических структур привела к образованию эукариотов я разделила их на грибы, простейшие, растительные и животные царства. По-видимому, разделение эукариотов на два царства (растения и животные) произошло еще до того, как образовались многоклеточные организмы, причем основным отличием животных и растений явилось наличие фотосинтеза у последних и отсутствие его у первых. Однако некоторые формы одноклеточных организмов трудно с определенностью отнести к одному из двух основных царств — животным или растениям. Таковыми являются некоторые формы одноклеточных, имеющих жгутики, например, Euglenavtrian и вообще все Protozoa. Вероятно, они также ранее отделились от общей ветви эукариотов, прежде чем произошла дивергенция растений и животных.
Дальнейшая эволюция органического мира происходила значительно более быстрыми темпами. К началу кембрия (570 млн лет назад) развились основные типы растений океанов и морей — водоросли. В результате их жизнедеятельности менялся состав атмосферы, которая из восстановительной становилась окислительной, приближаясь по своему составу к современной — с содержанием инертного озона, кислорода и небольших количеств углекислого газа, а также с водяными парами разной степени насыщенности. В течение первых двух периодов палеозоя началось проникновение растительных организмов на сушу — лишайники. Однако настоящее завоевание суши произошло в силуре (около 500 млн лет назад), когда появились псилофиты — предшественники мхов, плаунов, хвощей и папоротников. Выход растений на сушу сопровождался окончательным преобразованием атмосферы Земли, которая по своему составу стала близкой к современной. К этому же времени образовался слой озона в атмосфере, защищающий биосферу от значительной части ультрафиолетового излучения. В девоне (около 400 млн лет назад) и последующих периодах палеозоя и мезозоя развились многочисленные виды голосеменных, а в конце мезозоя (250 млн лет назад) появились покрытосеменные — современные деревья, кустарники и травы, которые вместе с хвойными стали преобладающими классами растений суши.
Эволюцию растений датируют с протерозойской эры (1,5 млрд лет назад), когда появились первые примитивные водные растения (водоросли), а также грибы. В палеозое (585—225 млн лет назад) получают развитие многоклеточные растения. В кембрии (585 млн лет назад) появляются многочисленные морские водоросли, в ордовике (505 млн лет назад) — наземные растения, которые получают дальнейшее развитие в силуре (425 млн лет назад). В девоне (375 млн лет назад) на суше, возникают леса, первые голосеменные растения. В карбоне (325 млн лет назад) господствуют папоротники, плауны, которые приходят в «упадок» в пермском периоде (240 млн лет назад). В мезозое (225—75 млн лет назад) отмечается господство голосеменных, особенно в триасе (225 млн лет назад) и юрском периоде (165 млн лет назад). Покрытосеменные появляются в юрском периоде и распространяются в меловом периоде (135'млн лет назад), полное «господство» их наступает в кайнозое (75 млн лет назад и до настоящего времени).
Покрытосеменные произошли от мезозойских голосеменных. По-видимому, однако, гингковые, хвойные, кордоиновые, пентоксилеевые, некоторые цикадовые и беннеттитовые, птеридоспермы и гнетавые не могут быть их предками, так как примитивные цветковые имеют лестничные трахеиды, а эти голосеменные — более высокоорганизованные, точечные. Кроме того, цветок предка должен быть обоеполым, а у них стробилы Однополые, и все названные группы голосеменных являются слишком специализированными формами. Наиболее древние остатки цветковых растений обнаружены в отложениях начала мелового периода, однако уже в это время они разнообразные и поэтому возникновение их следует отнести к более ранним эпохам. По мнению ряда авторов, покрытосеменные происходят не от единого предка — голосеменного, а от нескольких групп, в которых «рассеяны» признаки, характерные для цветковых. По-видимому, наряду с параллельной эволюцией большую роль играли ретикуляция эволюционных линий, обмен генетическим материалом в результате эпизодической гибридизации или вирусной трансдукции. В дальнейшем общее направление эволюции шло от древесных форм к кустарникам, затем к многолетним и, наконец, однолетним травам. Цветок покрытосеменных возник из энтомофильного стробила голосеменных. При этом решающую роль в эволюции цветка сыграли насекомые-опылители. Сначала появились двуполые цветки, позже — однополые, причем вначале средством для привлечения насекомых была пыльца, а затем нектар. Эволюция шла от спиральных цветков к циклическим, от полимерных к олигомерным, от одиночных к соцветиям. В настоящее время цветковые, или покрытосеменные, растения превалируют.
В растительном мире суши имеется до 300 000 видов покрытосеменных и колоссальное число их особей. За исключением лишайниковой и моховой тундры, сфагновых болот и хвойных лесов, занимающих довольно обширные пространства, суша покрыта главным образом представителями покрытосеменных. Форма покрытосеменных достигла необычайного разнообразия: от крошечной ряски до гигантских деревьев, от зеленых растений до сапрофитов, эпифитов и паразитов, от лиан до плотоядных росянок и мухоловок. По уровню организации покрытосеменные занимают такое же место в растительном мире, как млекопитающие в животном мире. Покрытосеменные представляют собой группу растений, приспособленных к наиболее полному использованию солнечной энергии.
Предков покрытосеменных растений следует искать среди голосеменных, однако не современных, высокоопециализированных форм, а среди более примитивных предков, например, семенных папоротников, беннеттитовых и цикадовых. Первоначальные покрытосеменные имели обоеполые цветки, которые развились из обоеполых стробил (шишек). Таким образом, предками покрытосеменных были примитивные группы семенных папоротников с трахоидами, имеющими еще лестничные утолщения и со свободными еще микроспоралиями.
Было высказано предположение о происхождении покрытосеменных от беннеттитов путем переноса признаков с одного пола на другой — гаметогетеротопии [Мейнен С. В., 1986]. У млекопитающих, птиц и насекомых этот феномен не столь уж редок, он приводит к скачкообразному изменению вида (например, уподобление женских, половых органов самок пятнистой гиены таковым самцов), поэтому данный феномен не может быть неожиданным у растений. Согласно этой гипотезе, гинецея покрытосеменных возникла от полиапермов беннеттитов. Вероятно, это были преимущественно горные формы тропических и субтропических стран, находившихся далеко от областей накопления осадков. Поэтому они не сохранились в виде ископаемых. Эволюция их происходила быстро в условиях неотении.
Большую роль в эволюции покрытосеменных сыграло развитие цветка, в образовании и эволюции которого важное значение имели насекомые. Цветок покрытосеменных возник из энтомофильного и обоеполого стробила. Насекомые, поедая пыльцу, способствовали перекрестному опылению. В то же время возникновение плодолистиков обеспечило защиту семе- зачатков и способствовало в дальнейшем появлению рыльца с функцией улавливания пыльцы. Все эти изменения имели исключительно большое значение в совершенствовании способов распространения семян. Древнейшие покрытосеменные были древесными растениями, но в дальнейшем появились кустарники, а затем травы—сначала многолетние, а позже однолетние. Эволюция также шла от вечнозеленых растений к растениям с сезонным появлением и опаданием листьев. Последние формы смогли распространиться в зоне умеренного и холодного климата.
«Обломками» древних покрытосеменных, живыми ископаемыми, сохранившимися до нашего времени, являются магнолиевые вечнозеленые деревья и кустарники, а также лавровые и троходендралы. Дальнейшая эволюция привела к разделению цветковых растений на одно- и двудольные, причем однодольные, вероятно, ведут свое происхождение от примитивных двудольных. Характерно, что среди однодольных отсутствуют истинные древесные формы: пальмы и другие однодольные древовидные формы не являются истинными деревьями и происходят от травянистых растений.
Покрытосеменные широко распространились в меловом периоде, когда происходили интенсивное горообразование и резкие изменения климата Земли. В этих условиях покрытосеменные обнаружили высокую эволюционную пластичность и необычайную приспособляемость, что и обеспечило быстрое расселение их по земной поверхности.
Выход жизни на сушу создал более благоприятные условия для накопления биомассы. Количество живого вещества суши в 200 раз превышает таковое живого вещества океанов, а выраженное в сухом виде — даже в 350 раз. При сравнении средних значений биомассы на единицу площади оказывается, что концентрация живого вещества на суше в 1000 раз больше, чем в океане.
Эволюция животных также начинается в протерозое, когда появились морские простейшие, а в конце эры — моллюски, черви и другие морские беспозвоночные. В кембрии господствуют трилобиты и плеченогие, зарождается большинство типов современных животных. Рыбы появляются в ордовике, но господствуют в море моллюски, кораллы и трилобиты. В силуре усиливается развитие рыб, господствуют морские наукообразные, появляются первые бескрылые насекомые. В девоне возникают земноводные, в карбоне появляются множество насекомых и первые пресмыкающиеся. Последние достигают расцвета в начале мезозоя (триас) и вымирают в конце мезозоя (меловой период). Кайнозойская эра характеризуется расцветом млекопитающих и птиц.
Простейшие имеют следующие 5 типов организации: моноэргидные (амебы), с полиплоидным ядром (радиолярии), полиэнергидные (полимастигиды), колониальные (уроглена), многоклеточные (вольвокс). Однако и многоклеточные простейшие по степени интеграции стоят значительно ниже самых примитивных метазоа.
К началу кембрия в водах мирового океана жили многие представители низших беспозвоночных — губки, медузы, кораллы, низшие черви, немертины. Последние уже имели системный уровень организации, ставши предшественниками (непрямыми!) высших животных. Дальнейшая эволюция животных шла в нескольких направлениях, давши ряд самостоятельных ответвлений — членистоногих, моллюсков, плеченогих и мшанок, иглокожих и др. Большинство из них появилось в ранних периодах палеозоя (кембрий, ордовик). Природа как бы нащупывала методом проб й ошибок возможные пути эволюции, неоднократно заходя в тупик. Большинство этих форм остались водными животными, которые не смогли достигнуть высоких стадий развития. Наиболее развитые моллюски — кальмары и осьминоги — имеют хорошо развитые органы чувств, однако их эволюция, длившаяся сотни миллионов лет, не привела к появлению зачатков разума. Наиболее процветающая группа высших беспозвоночных — членистоногие, особенно класс насекомых, стали заселять сушу в силуре (около 500 млн лет назад), после того как на суше появились растения (мхи и папоротники). За время длительной эволюции они дошли до стадии общественных насекомых (муравьи, пчелы, термиты) с высокоразвитыми инстинктами. Однако наличие внешнего скелета ограничило рост размеров тела, и эволюция их также не привела к появлению зачатков разума.
Эволюционная ветвь, которая привела к появлению позвоночных, также изобилует тупиковыми ответвлениями. Первые хордовые появились в силуре (490—420 млн лет назад). Сравнительно быстро эволюция их прошла от круглоротых до пресмыкающихся (330 млн лет назад), а затем снова «застопорилась» на 200 млн лет: в течение 120 млн лет на Земле господствовали динозавры, исчезнувшие в конце мелового периода, и только около 100 млн лет назад появились млекопитающие. В течение последних 70 млн лет от первичных млекопитающих образовались все ныне существующие формы плацентарных млекопитающих, и за этот же период возникли и вымерли многие промежуточные формы и их предшественники.
До сих пор во многом неясными остаются пути эволюции беспозвоночных, образовавших многочисленные таксономические группы высшего типа. Предполагают, что начальной формой многоклеточных был организм, сходный с паренхимулой Мечникова, обитавшей в воде. Он был покрыт экзодермальным ресничным эпителием и имел фагоцитобластическую паренхиму. В дальнейшем эволюция протекала в три этапа. На первом этапе образовались организмы, имевшие рот, который вел в фагоцитобластическую паренхиму; они напоминают бескишечных турбеллярий. Уже в это время организм стал питаться в целом, а не его клетки и в связи с этим в эктодерме появилось эпителиальное нервное сплетение. На втором этапе возникла эпителизированная кишка — второй энтодермальный эпителий. Формы, имевшие эпителизированный центральный фагоцитобласт, уже стояли на уровне кишечнополостных с более развитыми нервными элементами, формировавшимися за счет энтодермы. На третьем этапе организмы были сходны с греневиками. Они имели правильно разветвленную сеть гастроваскулярных каналов, заметно усложнился аборальный орган чувств.
Таким образом, эволюция исходных многоклеточных в толще воды могла привести к организации типа кишечнополостных с двумя отделами нервного аппарата и первичным аборальным органом чувств. Из них развились многочисленные другие формы беспозвоночных. Часть из них оседала на дне. Оседавшие на аборальный полюс дали начало Trochozoa, Acłinotrochozoa и Platyctenida; оседавшие на антимеру дали начало Scolecida, Brachiopoin, Chaetognata и Hemichoedata. У предков Scolecida (т. e. плоских и круглых червей и немертин) эволюция шла в направлении перемещения рта сначала на брюшную сторону, а потом по брюшной стороне в сагиттальной плоскости вперед вплоть до переднего терминального положения. Другое направление эволюции сколецид — появление эпителизированной кишки, а потом анального отверстия; при этом у них не образовалось энтодермальных нервных клеток и нервный аппарат возник из эктодермы. Дальнейшая эволюция этой ветви привела к появлению Hemichoedata. Из ветви, осевшей на оральный полюс, только Kamptozoa стали вести на дне прикрепленный образ жизни, оставшись первично бесцеломными; большинство же трохофоров стали вести подвижный образ жизни на дне. От них появились полихеты, моллюски, членистоногие. Из ветви, осевшей на аборальный полюс, развились иглокожие, губки, кишечнополостные. Эта группа наименее продвинулась эволюционно.
Предками позвоночных явились, по-видимому, первичные хордовые [Северцов А. Н., 1939], примитивные и неспециализированные многоклеточные беспозвоночные организмы, относящиеся к классу крыложаберных (Pterobranchia). Они обитали в кембрии (около 500 млн лет назад). Из них развились содержащие жаберные щели органулы, сходные с оболочечниками (Ascidiae), вернее, с их свободно плавающими личинками (ордовик, 420—350 млн лет назад), а от них произошли животные, напоминающие современного ланцетника. К концу силура (320 млн лет назад) появляются бесчелюстные (Agnatha) — рыбообразные существа (древние Ostracodermi). В позднем силуре и раннем девоне (320—300 млн лет назад) возникли первые челюстные позвоночные (Placodermi), а затем костные рыбы (Oxteichtys). В середине и конце девона (300— 280 млн лет назад) дивергировали кистеперые (Actinopterygii) и лопастеперые (Choaichtys) рыбы. Первые впоследствии образовали класс костистых рыб, вторые — немногие виды двоякодышащих рыб (Dipnoi) и латимерий. От них также ведут свое происхождение амфибии. Первые амфибии обнаружены в верхнем девоне (280 млн лет назад), они являются промежуточными формами между кистеперыми рыбами и типичными амфибиями. От этих примитивных форм неспециализированных амфибий ведут свое происхождение первые рептилии — стилозавры, появившиеся в -карбоне. Тем временем амфибии разделились на две ветви, давшие начало нынешним хвостатым, а также безногим, бесхвостым амфибиям.
Впервые сушу заселили амфибии, однако они остались тесно связанными в своем развитии с водой, тогда как рептилии приобрели приспособления к развитию яйца вне водной среды (амнион, аллантоис, большое количество желтка для питания зародыша, твердая скорлупа) и поэтому стали свободно расселяться на суше. Мезозойская эра (200—70 млн лет назад) была веком расцвета рептилий, из 5 подклассов рептилий Didpsida дали начало динозаврам, крокодилам, ящерицам и змеям, а также птерозаврам и птицам, a Synapsida — млекопитающим. Два последних класса позвоночных появились в конце юрского и начале мелового периода (130 млн лет назад). Среди млекопитающих три подкласса — однопроходные (Pro- totheria), сумчатые (Metatheria) и плацентарные (Eutheria) — представляют самостоятельные эволюционные ветви. Причем эти ветви, появившись 60—70 млн лет назад, стали особенно бурно развиваться в середине кайнозойской эры (около 20 млн лет назад). Наиболее древними и примитивными млекопитающими были насекомоядные, от них произошли ныне существующие и вымершие отряды плацентарных, включая приматов, появившихся в палеоцене (70—50 млн лет назад) [Simpson G., 1953]. Антропоиды появились в раннем олигоцене (35 млн лет назад), гоминиды — в раннем миоцене (25 млн лет назад), предки человека — в раннем плиоцене (10 млн лет назад). Около 1 млн лет назад появились древние люди —питекантропы и несколько позже неандертальцы. Последние вымерли во время четвертого ледникового периода и после этого оледенения около 50 000 лет назад появился современный человек.
В течение эволюции беспозвоночных можно видеть образование многочисленных тупиковых ветвей, не давших начало дальнейшей прогрессивной эволюции и тем более появлению разума. Такими своеобразными тупиковыми ветвями стали осьминоги и колониальные насекомые (муравьи, пчелы, термиты). Тупиковые ветви прослеживаются и среди позвоночных. Одним из таких тупиков эволюции явились динозавры. Они появились более 200 млн лет назад и существовали на Земле около 140 млн лет, исчезнув около 65 млн лет назад.
В заключение приведем календарь, дающий наглядное представление о масштабе времени эволюции органического мира [Волькенштейн М. В., 1984]. Вся эволюция жизни на Земле, если время существования Вселенной исчислять одним годом (масштаб 1 : 2 х 1012), на этой шкале заняла последний месяц года.
Начало года |
Большой взрыв |
Июнь |
Возникновение галактик |
Сентябрь |
Возникновение Солнечной системы и образование планеты Земля |
Октябрь |
Первые живые существа, древнейшие известные осадочные породы и окаменелые отпечатки микроорганизмов |
Ноябрь |
Микробионты, производящие кислород, широко развиваются. Возникновение полового способа размножения. Появление фотосинтезирующих растений, первые клетки, содержащие ядра (эукариоты) |
Начало декабря |
Образование кислородной атмосферы, интенсивные извержения вулканов. Развитие мейоза и полового способа размножения |
Середина декабря |
Развитие гетеротрофных одноклеточных, первые многоклеточные организмы. Начало макроскопической жизни |
20.ХІІ |
Возникновение беспозвоночных |
21.ХН |
Первый океанический планктон, расцвет трилобитов |
22.ХII |
Период ордовика; первые позвоночные (рыбы) |
23.ХІІ |
Силур; споровые растения завоевывают сушу |
24.ХІІ |
Девон; первые насекомые. Животные завоевывают сушу, первые амфибии, летающие насекомые |
25.ХII |
Каменноугольный период (каборн); первые хвойные растения, первые рептилии |
26.ХІІ |
Пермь; первые динозавры |
27.ХІІ |
Триас; первые млекопитающие |
28.XII |
Юра; первые птицы |
29.XII |
Меловой период; первые цветковые растения, вымирание динозавров |
30.ХII |
Третичный период; первые приматы, «расцвет» млекопитающих, первые гоминиды |
31.XII Около 14.00.00 ч |
Возникновение проконсула и рамапитека |
Около 22.30.00 ч |
Первые люди |
Около 23.00.00 ч |
Орудия из камня |
Около 23.59.00 ч |
Открытие земледелия |
Около 23.59.30 ч |
Первые города |
Около 23.59.54 ч |
Открытие письма |
Около 23.59.56 ч |
Бронзовая металлургия |
Около 23.59.57 ч |
Железная металлургия |
Около 23.59.59 ч |
Евклидова геометрия, архимедова физика |
24.00.00 ч |
Исчисление времени |
01.01 (Новый год) Около 00.00.01 ч |
Введение нуля и десятичного счета |
Около 00.00.02 ч |
Возрождение и современная наука |
Около 00.00.03 ч |
Современность |
Мы хотели бы заключить эту главу некоторыми соображениями об. эволюции генетического аппарата. Вначале он был невелик, и пока его молекулярная масса не превышала 106 (3000 пар нуклеотидов или сходного порядка величин), матричный синтез ДНК был достаточно надежен. С увеличением размера генома возрастала вероятность ошибок в процессе репликации, что стало непреодолимым препятствием для дальнейшего увеличения размеров генома. Вероятно, на этой стадии и появились «страховочные» приспособления, которые дошли до настоящего времени в виде системы Оказаки. Ее суть заключается в синтезе относительно небольших фрагментов-ДНК (до 3000 пар нуклеотидов), проверке правильности синтезов, исправлении ошибки и соединении фрагментов в непрерывную дупликационную нить. Эта система существует у всех современных прокариотов и более сложный ее аналог — у эукариотов. У вирусов (ДНК-содержащих) данная система может существовать или не существовать. В последнем случае синтез ДНК, по-видимому, происходит на более примитивной основе. Система Оказаки в то же время оказалась весьма громоздкой, и потребовалось усовершенствование работы генома в сторону большей его подвижности и оперативности. Таким «усовершенствованием» явились транспозоны и другие мигрирующие генетические элементы. Они также впервые появились у прокариотов, сохранились и получили дальнейшее развитие у эукариотов. У высших эукариотов (животных) возникла система интерферона, обеспечивающая гомеостаз нуклеиновых кислот. Эта система еще отсутствует у беспозвоночных и впервые появляется у хордовых, получив наивысшее развитие у млекопитающих и птиц. Она «сомкнулась» с иммунной системой, обеспечивающей белковый гомеостаз.