Зоология позвоночных: Учебное пособие - Т. А. Дауда 2014
Многоклеточные
Инфракласс Высшие звери, или плацентарные
Отряд Приматы
Отряд приматов объединяет около 200 видов исключительно разнообразных по внешнему виду млекопитающих животных (рис. 68). Их тело длиной от 9-12 см до 2 м, но, несмотря на внешние различия, их объединяют многие общие черты строения. Так, черепная коробка относительно велика, глазницы почти всегда направлены вперед, большой палец противопоставлен остальным, на пальцах ногти.

Рис. 68 Приматы:
1 — долгопят; 2 — черный ревун; 3 — павиан-бабуин; 4 — шимпанзе.
Задние лапы стопоходящие, а передние — хватательные. Соски (от 1 до 3 пар) располагаются на груди. Имеются все группы зубов, причем резцов две пары. В отряде два подотряда: Полуобезьяны и Обезьяны. Их представители обитают в тропических и субтропических странах.
Подотряд Полуобезьяны, или Низшие приматы. Полуобезьяны включают в себя приматов длиной от 9 до 106 см. У одних хвост длинный, густоопушенный, но не хватательный, у других — почти не опушенный или отсутствует. Самые примитивные среди них — тупайи, несколько похожие на белку, а по строению близки к наиболее древним насекомоядным млекопитающим. Более совершенную организацию, чем у тупайи, имеют лемуры и долгопяты. Их представители ведут древесный образ жизни, активны преимущественно ночью. Типичные виды лемуров — лемур вари и тонкий лори, а долгопятов — долгопят.
Подотряд Обезьяны, или Высшие приматы. Обезьяны объединяют самых высокоорганизованных животных на нашей планете.
Головной мозг у них относительно крупный, полушария переднего мозга у большинства видов имеют многочисленные борозды и извилины. Хвост отсутствует или развит в разной степени. Большой палец противопоставлен остальным. Тело длиной от 15 до 200 см. Известно около 140 видов.
Наиболее интересны среди высших приматов мартышки и человекообразные обезьяны. Мартышки имеют развитые защечные мешки, седалищные мозоли, хвост обычно длинный. Многие из них ведут древесный образ жизни, но еду отыскивают и на земле. Это очень подвижные, шумные животные, кричат по любому поводу. Передвигаются по земле и по толстым ветвям на четырех ногах, опираясь о поверхность ладонями рук и всей подошвой задних ног. К мартышковым относят собственно мартышек, павианов и макак.
Человекообразные обезьяны включают высших представителей отряда. Мозговой череп у них развит особенно сильно. Полушария переднего мозга имеют сложные борозды и извилины. Хвоста и седалищных мозолей нет. Все человекообразные обезьяны живут в лесах, легко лазают по деревьям, а приспособления к передвижению по земле несовершенны. Ноги при ходьбе они ставят не на всю подошву, а только на наружный край ступни. Движению и поддержанию равновесия помогают длинные руки, которыми животное опирается о землю тыльной стороной согнутых пальцев. Живут одиночно или группами. Человекообразных обезьян известно 11 видов; среди них — орангутан, шимпанзе, горилла и несколько форм гиббонов.
К высшим приматам принадлежит и человек. Однако он качественно отличается от всех других животных: по развитию трудовой деятельности, членораздельной речи, общественной жизни и сознания.
Вопросы для самопроверки и контроля:
1. Какие черты строения характеризуют млекопитающих?
2. Какие места обитания занимают млекопитающие?
3. Какими способами осуществляется терморегуляция у млекопитающих?
4. Какие особенности размножения позволяют делить млекопитающих на яйцекладущих, сумчатых и плацентарных?
5. Какие черты строения и образа жизни характерны для представителей подклассов яйцекладущих, сумчатых и отрядов плацентарных (на нескольких примерах).
6. Каково значение млекопитающих в природе и жизни человека?
Происхождение и эволюция птиц и млекопитающих
Птицы и млекопитающие возникли независимо, от далеко отстоящих друг от друга групп пресмыкающихся, причем сначала — млекопитающие, а потом — птицы.
Млекопитающие обособились от примитивных пресмыкающихся — зверозубых ящеров, еще сохранивших некоторые признаки земноводных, в триасе (примерно 215 млн лет назад).
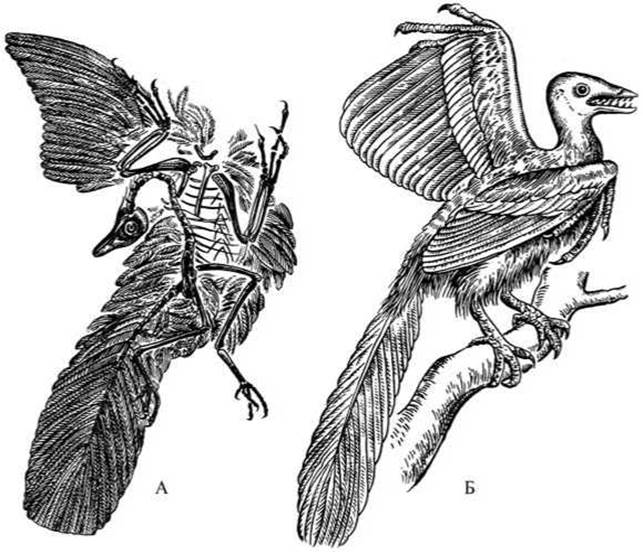
Рис. 69 Археоптерикс:
А — отпечаток на сланце; Б — реконструкция.
Обособление птиц от пресмыкающихся, вероятно, произошло 190-170 млн лет назад. Правда, ископаемых остатков древних птиц этого периода пока не найдено. В отложениях мелового залива юрского моря (возраст 150 млн лет) найдены отпечатки перьев и скелетов наиболее древней из известных птиц — археоптерикса, которого относят к особому подклассу ящерохвостых птиц (рис. 69). Предками современных — веерохвостых птиц, вероятно, были какие-то еще более примитивные и более древние ящерохвостые птицы.
Конкуренция и прямое преследование со стороны многочисленных и разнообразных пресмыкающихся вынуждали примитивных птиц и млекопитающих заселять наименее благоприятные, не освоенные или малоосвоенные биотопы, вырабатывать новые приспособления для добывания пищи, защиты от врагов, переживания неблагоприятных физико-химических воздействий.
В таких условиях наилучшим приспособлением, помимо перестроек в органах движения, пищеварении и других систем органов, было приобретение более гибкого поведения на основе усложнения центральной нервной системы и органов чувств.
В обоих классах возникли аналогичные, но развившиеся независимо приспособления, поднявшие жизнедеятельность птиц и млекопитающих на более высокий уровень по сравнению с другими позвоночными. В сложном комплексе таких приспособлений следует выделить:
✵ механизмы, поддерживающие устойчивую температуру тела при изменчивой температуре внешней среды;
✵ совершенствование нервной системы, органов чувств и усложнение поведения;
✵ расширение связей между особями и формирование на этой основе сложных форм внутривидовой организации, повышающей конкурентную способность вида в борьбе за существование.
Происхождение Хордовых
Ископаемые остатки хордовых не сохранились. Поэтому судить о ранних этапах их эволюции в значительной степени приходится по косвенным данным: путем сопоставления строения взрослых форм и сравнительного исследования эмбрионального развития.
Предков хордовых животных искали среди различных групп животных, в том числе и среди кольчатых червей. Предполагалось, что эти гипотетические предки перешли к активному образу жизни, но стали передвигаться на исходно спинной стороне тела. Предшественником хорды при этом мог бы стать соединительнотканный тяж, залегающий у некоторых полихет в толще вентральной мускулатуры (эти остроумные соображения подтверждения не получили).
Другая часть зоологов считает, что предшественниками хордовых, видимо, были целомические животные, перешедшие к малоподвижному образу жизни, что привело к уменьшению числа сегментов их тела и образованию вторичного рта. От таких животных обособились иглокожие, погонофоры — своеобразная группа животных-фильтраторов. Третья ветвь развития привела к обособлению хордовых животных. В самом начале от нее отделилась группа полухордовых, которой сейчас придают ранг типа. Обособление типа хордовых и его последующая эволюция связаны с увеличением подвижности, сопровождающейся общей интенсификацией жизнедеятельности и, прежде всего, совершенствованием добывания пищи.
Наиболее распространены две гипотезы происхождения хордовых.
По гипотезе Гарстанга (1928) предки хордовых, близкие к кишечнодышащим, перешли к сидячему образу жизни, усовершенствовав механизм фильтрации воды через прободенную жаберными отверстиями глотку. Подвижные личинки обеспечивали расселение. Такие личинки могли выработать способность к неотении — размножению на личиночной стадии, а взрослая фаза могла исчезнуть. Так мог возникнуть подвижный предок хордовых, подобно личинкам асцидий.
По гипотезе других исследователей (А. Н. Северцов, 1912, 1939; Н. А. Ливанов, 1958), хордовые могли произойти от червеобразных ползающих или роющих предков, по внешнему виду напоминающих современных кишечнодышащих, у которых развилась хорда, а в прободенной жаберной щелями глотке возник эндостиль — орган, выделяющий слизь и обеспечивающий улавливание пищи из фильтруемой воды. Это превратило глотку в мощный аппарат фильтрации, подобно глотке современного ланцетника.
Обе гипотезы требуют дальнейшей разработки. Они исключают друг друга, но обе признают, что у предков хордовых питание было пассивным. Такой тип питания связан с малоподвижным образом жизни. Из двух способов перехода к подвижному существованию и активному питанию — путем неотении или путем возникновения роющих животных — более вероятным кажется второй, так как личинки асцидий и неотенические личинки самостоятельно не питаются.
Хордовые в родословном древе животного мира
Изучение многочисленных данных зоологической науки подводит к материалистическому и диалектическому пониманию путей развития живой природы.
Всесторонние исследования убеждают в происхождении животных от общего корня, в монофилетическом развитии всего животного мира. Поэтому филогенетические отношения животных разных систематических групп могут быть изображены в виде разветвленного древа, в основании которого находятся примитивные одноклеточные животные (рис. 70).
Установлению филогении животных, выявлению их родственных взаимоотношений способствуют три пути исследования:
✵ сравнительное изучение морфологии современных животных (сравнительная морфология);
✵ изучение их эмбриологии также в сравнительном освещении (сравнительная эмбриология);
✵ изучение по ископаемым остаткам животных прошлых геологических эпох (сравнительная палеонтология).
Сравнительно-морфологический метод изучения животных позволяет вскрыть разную степень сложности их строения и обнаружить те или иные признаки сходства и различия, что помогает составить представления о возможных переходных формах и гипотетических предках той или иной группы животного мира.
Например, выяснено, что плоские гребневики — животные, близкие к кишечнополостным, в то же время имеют ряд черт сходства с ресничными червями из типа плоских червей, сочетая в строении признаки двухслойных животных с радиальной симметрией тела с признаками трехслойных животных с двусторонней симметрией. Подобное сходство позволяет высказать предположение о филогенетической связи плоских червей с кишечнополостными.
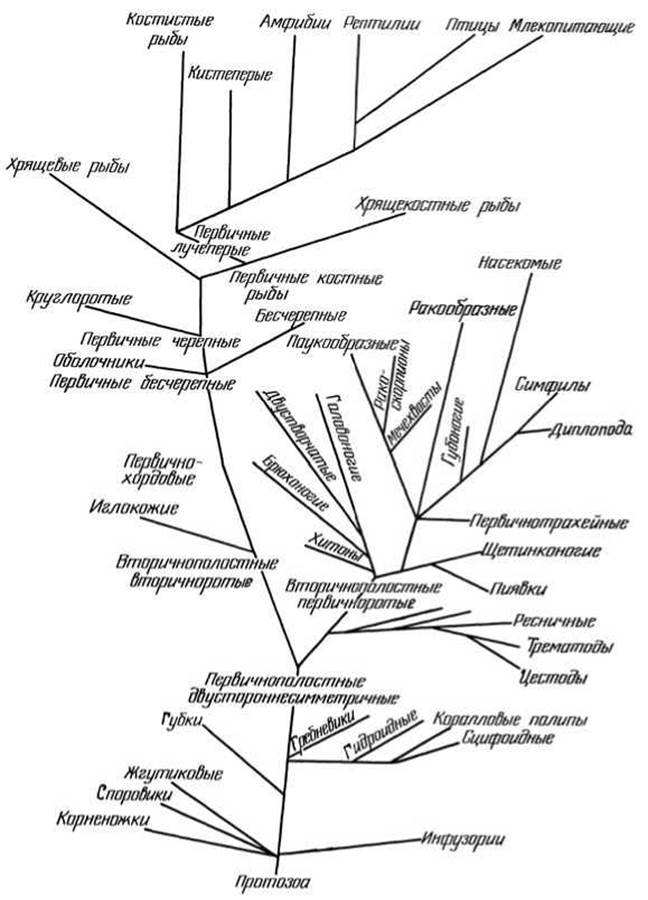
Рис. 70 Родословное древо животного мира (схема)
Использование данных сравнительной эмбриологии основывается на биогенетическом законе (правиле), согласно которому прошлое не проходит в эволюции животных бесследно и животные в индивидуальном развитии повторяют в той или иной степени этапы своего исторического развития — онтогенез отражает филогенез. Основанием для формулировки этого закона известным немецким дарвинистом Э. Геккелем послужили многочисленные исследования корифеев нашей отечественной науки А. О. Ковалевского и И. И. Мечникова — по эмбриологии беспозвоночных и низших хордовых и К. М. Бэра — по эмбриональному развитию позвоночных. Сходство в эмбриональном развитии животных может служить дополнительным аргументом при решении вопроса о филогенетических связях.
Палеонтологический путь исследования филогении животных также дает веские доказательства для установления родословной тех животных, остатки которых сохранились в слоях земли.
Одноклеточным организмам со всеми характерными для них чертами сложного строения — наличия цитоплазмы, ядра и различных органоидов, выполняющих жизненные отправления — предшествовали живые существа значительно более простой организации, неклеточного строения. Первые организмы обладали способностью воссоздать живую материю тела путем ассимиляции из окружающей среды неорганических и органических веществ небиологического происхождения. От них произошли одноклеточные животные, возможно, амебоидного строения.
Среди жгутиковых появились организмы, способные к построению тела из неорганических соединений с использованием солнечной энергии в процессе фотосинтеза. Зеленые жгутиковые, соединяющие в строении и биологии черты животных и растений, свидетельствуют о происхождении двух царств органической природы от общих предков.
Эволюция одноклеточных протекала в разных направлениях, но одноклеточная организация тела у большинства сохранялась. Древние амебоидные организмы дали начало современным саркодовым и паразитическим книдоспоридиям. Одна из ветвей родословного древа от жгутиковых идет к инфузориям — животным наиболее сложного строения, другая — к паразитическим споровикам, хотя о происхождении споровиков и книдоспоридий нет единого мнения.
Развитие одноклеточного животного мира привело через колониальных жгутиковых к многоклеточным животным. Жгутиковые отличаются среди Protozoa наибольшим разнообразием, сложностью приспособлений к питанию и жизни в разных условиях. Есть среди них простые и сложные организмы колониальной формы, давшие начало двум ветвям современных многоклеточных животных: губкам и остальным многоклеточным — через первичных двухслойных предков.
Существует несколько гипотез о пути происхождения многоклеточных, и все они объединены общим признанием происхождения многоклеточных от колониальных жгутиковых. От воротничковых жгутиковых произошли губки, неподвижный образ жизни которых не способствовал их прогрессивному развитию, поэтому они сохранили много примитивных черт строения. От других колониальных жгутиковых произошли двухслойные предки кишечнополостных животных. Большинство их перешло к неподвижному или малоподвижному существованию, как это наблюдается у современных кишечнополостных.
Сравнительное изучение двухслойных кишечнополостных приводит к представлению о возможных путях происхождения и эволюции тканей, дифференцировки их функций и специализации. Кишечнополостные имеют в мезоглее клеточные структуры различной сложности и назначения, которые помогают представить этапы развития третьего зародышевого листка — мезодермы. Зачатки мезодермы, имеющиеся у кишечнополостных, еще более отчетливо выражены у гребневиков — животных, по происхождению и организации близких к кишечнополостным. Вопрос о происхождении плоских червей продолжает оставаться дискуссионным. Гипотеза Ланга о филогенетической близости их с плоскими ползающими гребневиками оспаривается, но ряд новых гипотез также не решает вопроса. Ланг основывает свою гипотезу на морфологическом сходстве гребневиков и ресничных червей. Последние, подобно некоторым плоским гребневикам, имеют уплощенное тело, покрытое ресничным эпителием, нервную систему в виде сети нервных клеток или собранных в нервных тяжах; кишечный канал тех и других лишен анального отверстия. У некоторых турбеллярий наряду с двусторонней симметрией тела сохраняются черты радиальной симметрии в расположении отдельных органов: центральное положение рта на брюшной стороне, радиальное расхождение от глотки ветвей кишечника, расположение нервного узла почти над глоткой с радиально расходящимися от него нервными тяжами.
В родстве с плоскими червями состоят круглые черви и скребни. Тип Круглые черви включает класс Брюхоресничные, представители которого имеют как во взрослом состоянии, так и в период эмбрионального развития ряд сходных черт строения с ресничными плоскими червями (наличие ресничного эпителия на брюшной стороне, типичных протонефридиев и др.). Это дает основание сближать круглых червей с плоскими.
От предков современных первичнополостных отделились вторичнополостные животные (целомата), которых с полным основанием называют высшими многоклеточными животными. Основанием этому служат такие черты их организации, как концентрации нервных клеток в ганглиях — прогрессивное развитие нервной системы, образование кровеносной сосудистой системы, выделительных органов метанефридиального типа, сегментарность тела.
Одну из ветвей вторичнополостных животных составляют организмы, у которых во взрослом состоянии сохраняется первичный рот (первичноротые). Раньше других от первичных вторичнополостных животных отделилась ветвь, приведшая к возникновению типа моллюсков. Несмотря на то что другие моллюски имеют на всех этапах онтогенеза несегментированное тело, родство их с другими вторичнополостными животными не вызывает сомнения. Как у тех, так и других имеются нервные ганглии, кровеносная система, метанефридиальные выделительные органы. Такое глубокое сходство организации не может быть случайным. Кроме того, типичная для морских моллюсков личинка — парусник — сходна с личинкой многощетинковых кольчатых червей трохофорой.
Среди сегментированных вторичнополостных животных исходной группой следует считать древних первичных многощетинковых червей, близких к типу Кольчатые черви. Они дали начало малощетинковым червям, а от последних произошли пиявки.
Сравнение кольчатых червей с членистоногими убеждает в большом сходстве их строения: у тех и других имеется вторичная полость тела, метамерность, нервная система в виде надглоточных ганглиев, окологлоточного кольца и брюшной нервной цепочки, кровеносная система с главным сосудом на спинной стороне и другие общие признаки; посегментное расположение конечностей, которые у многощетинковых кольчатых червей имеют вид нерасчлененных параподий, а у членистоногих вместе с приобретением твердого покрова преобразовались и стали членистыми ножками. Общие черты строения кольчатых червей и членистоногих указывают на близкое родство тех и других. Членистоногие — прогрессивная ветвь, берущая начало от общего предка с современными многощетинковыми кольчатыми червями.
Среди членистоногих ракообразные, продолжающие жить в воде, где жили и их предки, сохранили многие примитивные признаки строения, сближающие их с кольчатыми червями.
Паукообразные во многом сходны с ракообразными, ведут происхождение от древних жабродышащих членистоногих. В родстве с предками — кольчатыми червями — находятся и трахейнодышащие членистоногие, среди которых наиболее прогрессивной группой являются насекомые.
Другая ветвь родословного древа развития животного мира ведет от древних вторичнополостных червеобразных животных к вторичноротым животным — иглокожим и хордовым.
Иглокожие — древняя группа животных, которые вторично приобрели радиальную симметрию тела. Она появилась у палеозойских представителей этого типа в связи с переходом к прикрепленному образу жизни. Личинки иглокожих имеют двустороннесимметричное тело.
Низших хордовых с иглокожими сближает вторичное образование рта в процессе онтогенеза, сходство процесса образования вторичной полости тела, двухслойное строение покровов и ряд других признаков.
Хордовые животные — одна из наиболее высокоорганизованных групп животного мира. Происхождение хордовых остается в деталях еще неясным. Возможно, они произошли от общего корня с иглокожими.
Первичные хордовые, как показали исследования А. Н. Северцова, представляли собой существа, довольно близкие к современному ланцетнику. Это были вторичнополостные, вторичноротые животные с признаками внутренней сегментации, имевшие хорду, нервную трубку вдоль спины и жаберные щели в стенках глотки. От этих примитивных первичных хордовых произошли, с одной стороны, современные бесчерепные (ланцетники и близкие к ним формы), а с другой — первичные черепные (позвоночные), у которых передняя часть спинного мозга преобразовалась в головной мозг и защищать его стал сформировавшийся череп. Боковую ветвь низших хордовых образуют личиночнохордовые, в организации взрослых особей которых произошел регресс в связи с переходом к сидячему образу жизни при пассивном питании.
В эволюции позвоночных одна из ветвей привела к появлению бесчелюстных (миноги и миксины), а другая дала начало челюстноротым, к которым относятся все остальные классы Vertebrata. У животных этой ветви в связи с активным добыванием пищи и общим нарастанием активности поведения развились парные конечности, челюстной аппарат, сложный скелет и ряд других усложнений в строении. Все это обеспечивало им развитие и широкое распространение. Первыми представителями челюстноротых позвоночных были рыбы, в первую очередь хрящевые (акулы и скаты). Но уже в конце палеозоя широкое распространение получают различные рыбы с костным или костно-хрящевым скелетом. От древних палеозойских рыб ведут родословную двоякодышаще и кистеперые рыбы. Последних надо считать родоначальниками наземных позвоночных, древнейшими из которых были панцирноголовые земноводные — стегоцефалы. От древних примитивных амфибий произошли как земноводные, так и первые палеозойские пресмыкающиеся, достигшие в мезозойскую эру большого разнообразия форм.
В мезозое от пресмыкающихся отделились два высших класса позвоночных: Млекопитающие и Птицы. Как уже было сказано, в юрских отложениях найдены остатки своеобразных существ — археоптериксов, которые совмещали признаки рептилий и птиц. Это боковая ветвь в эволюции птиц. Настоящие птицы появились в меловом периоде и достигли большого разнообразия в палеогене и неогене кайнозойской эры.
Млекопитающие произошли от древних рептилий в самом начале мезозойской эры, т. е. раньше птиц. Но развитие, приведшее к современному богатству форм этого класса позвоночных, относится, как и у птиц, к кайнозойской эре.