ЗООЛОГІЯ БЕЗХРЕБЕТНИХ - Г. Й. Щербак - 2008
ЦАРСТВО БАГАТОКЛІТИННІ (METAZOA)
Багатоклітинні опанували всі основні середовища існування - водне, наземно повітряне, ґрунтове та організми інших істот. Нараховують понад 1,5 млн видів.
Тіло багатоклітинних складається з багатьох клітин, які, на відміну від клітин колоній найпростіших, розрізняються як за будовою, так і за функціями. У переважної більшості багатоклітинних подібні клітини зібрані у тканини, з яких складаються органи та системи органів.
Багатоклітинним тваринам властиве нестатеве та статеве розмноження, проте переважаючою формою, а в деяких групах і єдиною, є статеве розмноження.
Нестатеве (вегетативне) розмноження багатоклітинних - це утворення нових організмів без участі статевих клітин. Воно може відбуватися різними способами: поділом (поперечним, поздовжнім або невпорядкованим), брунькуванням (зовнішнім або внутрішнім), лацераціею (розпадом материнського організму на кілька частин і відновленням з кожної частини цілого організму) тощо.
Статеве розмноження здійснюється за участі спеціальних генеративних (статевих) клітин. На відміну від решти клітин - соматичних, які в багатоклітинних є диплоїдними, гамети, що утворюються з генеративних клітин, є гаплоїдними, при цьому відбувається редукційний поділ (мейоз), тобто їхній ядерний цикл супроводжується гаметичною редукцією хромосом (рис. 1). Виняток становлять лише коловертки, які мають життєвий цикл з проміжною редукцією. Статевий процес у багатоклітинних є оогамним: яйцеклітини (макрогамети) та сперматозоїди (мікрогамети) відрізняються не лише за розмірами, а й за будовою. Яйцеклітини нерухомі, позбавлені джгутиків і мають переважно сферичну форму, часто вони містять жовток - гранули запасних поживних речовин, необхідних для розвитку зародка. В яйцеклітині розрізняють два полюси: анімальний, де міститься ядро й розташована зона інтенсивного обміну речовин, та протилежний - вегетативний. Типовий сперматозоїд має будову видозміненого джгутикового. Нерухомий сперматозоїд зветься спермієм. Після запліднення яйцеклітина вкривається однією або кількома оболонками й стає яйцем. Однією із форм статевого розмноження є партеногенез, за якого новий організм утворюється з незаплідненої яйцеклітини, яка в такому разі не зазнає редукційного поділу.
Життєвий цикл багатоклітинних характеризується складним індивідуальним розвитком - онтогенезом., у процесі якого із заплідненої (а в разі партеногенезу - з незаплідненої) яйцеклітини утворюється статевозрілий організм. Онтогенез складається з кількох етапів.
1. Спочатку відбувається гаметогенез, у результаті якого з недиференційованих диплоїдних клітин шляхом мейозу утворюються гаплоїдні яйцеклітини й сперматозоїди. Після запліднення яйцеклітини починається дроблення яйця. Типи дроблення бувають різними й залежать від кількості та розміщення жовтка в яйці: поєне й рівномірне з утворенням однакових клітин (бластомерів), якщо жовтка в яйці небагато й він розміщений рівномірно; нерівномірне - якщо жовток сконцентрований на вегетативному полюсі, тоді на цьому полюсі формуються великі, багаті на жовток макромери, а на анімальному - дрібні мікромери; якщо жовтка дуже багато, відбувається поверхневе дроблення, за якого бластомери розмішуються поверхнево, а центральна маса жовтка залишається неподіленою або бластомери утворюються лише на анімальному полюсі, а вегетативна частина яйця із жовтком не ділиться.
Типи дроблення також визначають за взаємним розташування бластомерів, що утворилися (рис. 54). Архаїчне дроблення - це, коли бластомери розташовані неупорядковано. Під час спірального дроблення мітотичні веретена розміщуються під кутом 45° до поздовжньої осі яйця за годинниковою стрілкою або проти, причому в кожному наступному поділі напрямки чергуються, і в результаті бластомери розташовуються по спіралі. За радіального дроблення перші дві борозни йдуть від полюса до полюса меридіонально, третя - по екватору. Потім меридіональні та екваторіальні борозни чергуються, і бластомери розмішуються правильними горизонтальними та вертикальними рядами. Зрідка має місце білатеральне дроблення, коли бластомери розподіляються симетрично до поздовжньої площини симетрії.
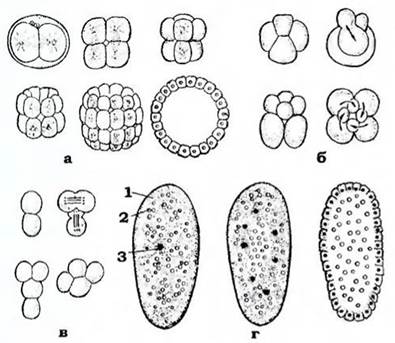
Рис. 54. Типи дроблення яйця
(з Іванової-Казас, зі змінами): а - радіальне; б - спіральне; в - білатеральне; г - поверхневе:
1 - цитоплазма; 2 - гранули жовтка; 3 — ядро
Дроблення завершується утворенням стадії бластули. Найчастіше вона має вигляд кулі, яку утворено поверхневим шаром клітин, усередині якого є більш-менш розвинена порожнина - бластоцель, заповнена рідиною. Іноді вона вся заповнена клітинами, і такий зародок зветься морулою.
2. Далі починаються процеси переміщення клітин - гаструляція, які приводять до утворення двох або трьох зародкових листків: ектодерми, ентодерми та мезодерми (рис. 55). Спочатку формується двошаровий зародок - гаструла. Вона складається з поверхневого шару клітин - ектодерми та внутрішнього - ентодерми, яка вистилає порожнину первинної кишки - гастроцель. Остання на вегетативному полюсі має отвір - первинний рот, або бластопор.
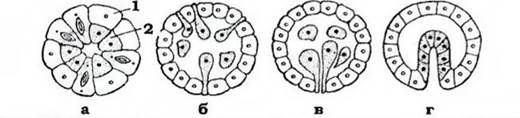
Рис. 55. Основні типи гаструляції (з Іванової- Казас, зі змінами): а - деламінація; б - мультиполярна імміграція; в - уніполярна імміграція; г- інвагінація: 1 - ектодерма; 2 - ентодерма
Розрізняють кілька типів гаструляції:
імміграція - частина клітин бластули мігрує у бластоцель, де утворює шар ентодерми. Імміграція буває мультиполярною, коли проходить по всій поверхні бластули, та уніполярною, якщо клітини мігрують лише з вегетативного полюса. При цьому бластопор часто не виражений;
деламінація - клітини бластули поділяються навпіл паралельно поверхні, утворюючи таким чином одразу екто- та ентодерму;
інвагінація - на вегетативному полюсі стінка бластули вгинається у бластоцель, і утворюється гаструла у вигляді двошарового мішка з бластопором і первинною кишкою; порожнина між екто- та ентодермою має назву бластоцель;
епіболія - великі клітини вегетативного полюсу обростають дрібними клітинами анімального, при цьому гастроцель не утворюється, а бластопор має вигляд заглиблення на вегетативному полюсі.
Способи утворення третього зародкового листка - мезодерми - є досить різноманітними й розглядатимуться у відповідних розділах.
3. Клітини зародкових листків диференціюються, утворюючи тканини та зачатки органів. З ектодерми формуються поверхневий шар шкіри та його похідні (шкірні залози, кутикула, черепашка тощо), нервова система та органи чуття, епітелій переднього та заднього відділів кишечнику, органи виділення (протонефридії, метанефридії, мальпігієві судини комах, багатоніжок), органи дихання безхребетних; з ентодерми - середній відділ кишечнику, травні залози, мальпігієві судини в павукоподібних; мезодерма дає початок паренхімі, статевим залозам (гонадам), целомічному епітелію, м'язам.
Ембріональний розвиток відбувається під покривом яйцевих оболонок або в тілі материнського організму.
4. Після виходу з яйця (або народження) починається постембріональний розвиток, який може бути прямим або з перетворенням - метаморфозом. За прямого розвитку з яйця виходить молодий організм, подібний до дорослого, він відрізняється від дорослого лише розмірами та недорозвиненими статевими органами. Розвиток з метаморфозом характеризується різноманітними личинковими стадіями, які відрізняються від дорослих особин будовою та способом життя. У багатьох безхребетних із зовнішнім скелетом або товстою кутикулою (нематоди, членистоногі) ріст личинок супроводжується линяннями.
Онтогенез завершується формуванням дорослої тварини, готової до статевого розмноження. Від цієї схеми відрізняється онтогенез первинних багатоклітинних (Prometazoa), які не мають зародкових листків.
Залежно від рівня організації царство Багатоклітинні поділяють на два підцарства: Первинні багатоклітинні (Prometazoa) та Справжні багатоклітинні (Eumetazoa).