ЗООЛОГІЯ БЕЗХРЕБЕТНИХ - Г. Й. Щербак - 2008
ЦАРСТВО БАГАТОКЛІТИННІ (METAZOA)
ПІДЦАРСТВО СПРАВЖНІ БАГАТОКЛІТИННІ (EUMETAZOA)
РОЗДІЛ ТРИШАРОВІ (TRIPLOBLASTICA), або БІЛАТЕРАЛЬНІ (BILATERIA) ТВАРИНИ
ПІДРОЗДІЛ СПІРАЛЬНІ (SPIRALIA)
ТИП ПЛОСКІ ЧЕРВИ (PLATHELMINTHES)
ПІДТИП НОВОШКІРІ (NEODERMATA)
НАДКЛАС ЦЕРКОМЕРНІ (CERCOMEROMORPHA)
КЛАС МОНОГЕНЕЇ (MONOGENEА)
Більшість видів моногеней - ектопаразити хребетних тварин; незначна кількість видів - ендопаразити ротової порожнини, задньої частини кишечнику, органів видільної (сечоводи риб, сечовий міхур жаб) чи кровоносної систем. Здебільшого паразитують на прісноводних і морських рибах, значно рідше - на амфібіях і рептиліях. І лише один вид Oculotremahippopotami є паразитом кон'юнктиви ока бегемотів. Клас налічує близько 2 тис. видів (в Україні - понад 200 видів).
Моногенеї, або моногенетичні сисуни, відрізняються наявністю органа прикріплення - церкомера з гачками або присосками, деякими деталями будови травної, видільної та статевої систем і простим життєвим циклом з метаморфозом, але без зміни хазяїв.
Будова. Тіло моногеней видовжене, листоподібне, від 0,02 мм до 3 см завдовжки. Воно поділене на власне тулуб і прикріпний диск - церкомер (опістаптор) із хітиноїдними гачками, ними паразит закріплюється у тканинах хазяїна (рис. 133). У більш спеціалізованих видів на церко- мері є ще й присоски або клапани. Рідше весь прикріпний диск перетворюється на один великий присосок, який вторинно може поділятися перегородками на окремі комірки (наприклад, у Nitzschia). Інколи церкомер набуває досить химерної форми, наприклад, У Heterobothrium affinis він має пальцеподібні вирости, що нагадує руку, яка ніби охоплює зяброві пелюстки хазяїна (рис. 133, в). На передньому кінці тіла можуть розмішуватися рухомі головні лопаті, на поверхні яких відкриваються протоки одноклітинних залоз з клейким секретом.
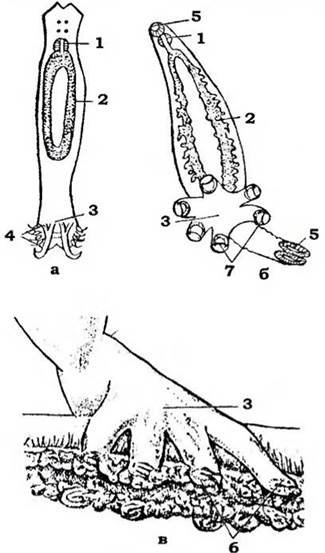
Рис. 133. Травна система та прикріпні апарати моногеней (із Стрелкова):
а - Dactylogyrus; б -Hexabothrium; в - Heterobothrium affinis:
1 - глотка; 2 - середня кишка; 3 - прикріпний диск; 4 - гачки; 5 - присоски; 6 - прикріпні клапани; 7 - серпоподібні гачки
Шкірно-м'язовий мішок. Під типовим тегументом залягають шари кільцевих і поздовжніх м'язів, іноді між ними лежить шар діагональних м'язів. З тегументом у деяких видів пов'язані численні шкірні залози, протоки яких відкриваються на поверхню тіла. їхні функції ще залишаються не з'ясованими.
Травна система подібна до такої у трематод. Середня кишка мішкоподібна або двогілляста, інколи у вигляді замкненого кільця чи з бічними виростами. У великих видів головні гілки кишечнику утворюють численні відгалуження у вигляді густої сітки.
Примітивні представники класу живляться слизом та епітеліальними клітинами хазяїна. При цьому види роду Gyrodactylus розривають епітелій хазяїна за допомогою виростів пірамідальних клітин глотки. Представники родини Polystomatidae - облігатні (обов'язкові) гематофаги. Залози передніх відділів їхнього кишечнику виділяють антикоагулянти, які запобігають згортанню крові хазяїна.
Видільна система протонефридіальна, представлена парними каналами, кожен з яких відкривається самостійним отвором.
Нервова система - типовий ортогон. Добре розвинена іннервація прикріпного диска.
Органи чуття розвинені слабо. На поверхні тіла розташовані сенсили - одиничні й зібрані у групи. Інвертовані очі здебільшого функціонують лише на личинковій стадії, у дорослих особин вони рудиментарні або зовсім відсутні.
Статева система моногеней гермафродитна. Чоловіча статева система представлена сім'яниками, сім’явивідними протоками, додатковими залозами та копулятивним апаратом, часто складної будови. Кількість сім'яників значно варіює. Деталі будови жіночої статевої системи у різних видів також можуть бути різними. Яєчник завжди один, жовтівники добре розвинені. Є піхва, через яку сперма партнера потрапляє до оотипу. Моногенеї відкладають складні яйця.
Розмноження. Запліднення зазвичай перехресне, але можливе й самозапліднення. Унікальне пристосування для забезпечення перехресного запліднення є у паразитів коропових риб з роду Diplozoon. Ще на стадії личинки дві особини зростаються між собою таким чином, що чоловіча статева протока (сім'япровід) однієї особини відкривається в жіночу протоку (піхву) іншої (рис. 134). Без цього вони не досягають статевої зрілості.
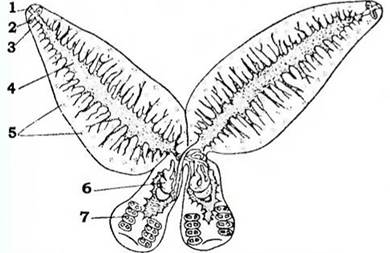
Рис. 134. Diplozoon paradoxum (із Стрелкона):
1 - рот; 2 - біляротові присоски; 3 - глотка; 4 - середня кишка; 5 - жовтівники; 6 - комплекс статевих органів; 7 - прикріпні клапани
Життєвий цикл моногеней, за незначними винятками, проходить без чергування поколінь і зміни хазяїв. Живородіння зустрічається рідко (наприклад, рід Gyrodactylus). Найбільш прості життєві цикли спостерігаються в моногеней роду Dactylogirus, в яких з яєць виходять личинки з поясками війок, двома парами очей і церкомером на задньому кінці тіла. Личинки прикріплюються до зябер риб і перетворюються на дорослих особин.
Складніший життєвий цикл у жаб’ячої багатоустки (Polystomum integerrimum) (рис. 135), для нього характерна циклічність, пов'язана з особливостями біології хазяїна. Статевозрілі особини паразитують у сечовому міхурі жаб і живляться кров'ю. Час відкладання яєць багатоустом збігається з періодом відкладання жабами ікри у воду. Личинки паразита, які вийшли з яєць, інвазують пуголовків жаб, які щойно вийшли з ікринок, але оселяються вони не в сечовому міхурі, а на їхніх зябрах. Особини ектопаразитичного покоління є значно дрібнішими, ніж ендопаразитичного, в них слабше розвинений церкомер, відсутня піхва. Вони досягають статевої зрілості на зябрах пуголовка й відкладають у воду незначну кількість яєць, після чого гинуть. Личинки, які виходять із цих яєць (наступне покоління), знову заражають пуголовків. Але на зябрах вони завершити свій розвиток уже не встигають - зябра пуголовків редукуються й зяброва порожнина заростає. Тому багатоусти мігрують поверхнею тіла пуголовка й через клоаку потрапляють до сечового міхура, де й завершується розвиток ендопаразитичного покоління. Зараження жаб відбувається лише раз упродовж їхнього життя, на стадії пуголовка. Живуть паразити до шести років (для порівняння: ектопаразити риб з родини Dactylogyridae - всього 25-40 діб).
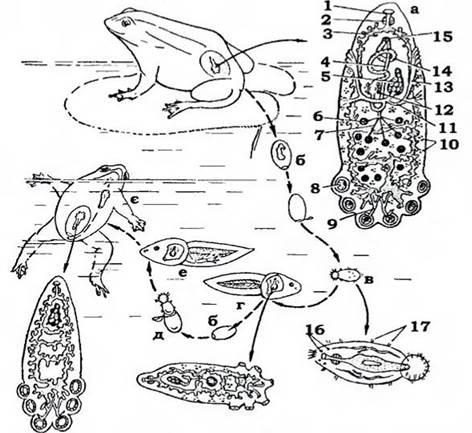
Рис. 135. Життєвий цикл Polystomum integerrimum:
а - паразит із сечового міхура жаби; б - яйце; в - личинка; г - зяброва форма паразита на пуголовку; д - вихід личинки з яйця;
е - пуголовок наприкінці метаморфозу з личинками другого покоління паразита; є - міграція личинки до сечового міхура жаби;
ж - незріла форма паразита із сечового міхура жаби:
1 - рот; 2 - глотка; 3 - середня кишка; 4 - сім'япровід; 5 - яєчник; 6 - сім'яник;
7 - сім’явиносний канал; 8 присосок; 9 - гачки; 10 - жовтівники;
11 - жовточні протоки; 12 - оотип, оточений тільцем Меліса; 13 - піхва;
14 - матка; 15 - статевий отвір; 16 - очі; 17 - пояски війок
Найбільшої шкоди моногенеї завдають ставковим рибним господарствам. Це, зокрема, представники родин Dactylogyridae та Gyrodactylidae. Найбільше від них страждають мальки й молодь риб. Руйнуючи епітелій зябер, паразити порушують газообмін і спричинюють запалення. У разі високої інтенсивності інвазії (до 500 паразитів на одну особину) дактилогіруси можуть викликати загибель риби, інколи масову.
Відомі випадки, коли моногенеї викликали масову загибель хазяїв й у природних екосистемах. Так, після спроби акліматизувати волзьку севрюгу в Аральському морі паразити цього виду осетрових риб - Nitzschia sturionis - перейшли до паразитування на місцевому аральському шипі, що викликало його масову загибель.