ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 2 - Г.Й. Щербак - 1996
ТИП ЧЛЕНИСТОНОГІ (ARTHROFODA)
ПІДТИП ТРАХЕЙНОДИШНІ (TRAСHEАТА)
КЛАС КОМАХИ, АБО ВІДКРИТОЩЕЛЕПНІ (INSECTA, АБО ECTOGNATHA)
Більшість видів комах — мешканці суші. Вони заселяють усі континенти, включаючи Антарктиду, і трапляються скрізь: у безводних пустелях, високогірних зонах вічних снігів, лісах і степах. Комахи освоїли всі типи наземних біоценозів, а також ґрунти. Чимало їх живе у прісних водоймах. Проте цілком водяними тваринами, що дихають розчиненим у воді киснем, стали тільки личинки комах, дорослі крилаті особини яких здатні залишати воду. Комахи в основному вільноживучі тварини, але серед них чимало і паразитів.
Кількість видів комах, що заселяють земну кулю, ще точно не встановлено: за різними підрахунками вже описано від одного до трьох мільйонів сучасних видів, і щорічно вчені відкривають кілька тисяч нових для науки видів. Видовий склад комах України вивчено ще недостатньо: вважають, що є не менше ніж 40 тис. видів.
Розміри комах коливаються у широких межах — від 0,25 мм до 26 см.
Тіло комах складається з трьох тагм: голови, грудей та черевця (рис. 118). Голова має ротові органи та одну пару вусиків. Груди складаються з трьох сегментів і несуть три пари ніг та, як правило, крила. Черевце складається з 11 сегментів і, як звичайно, не має ніг.
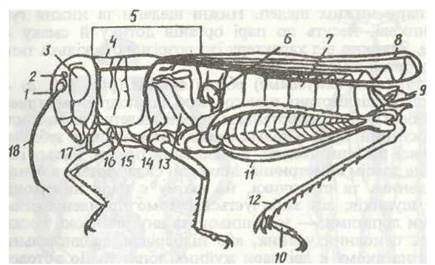
Рис. 118 Тіло комахи
Голова вкрита суцільною капсулою, вона утворилася в результаті злиття кількох сегментів та акрона. На ній розташовані пара складних (фасеткових) очей, прості вічка, пара вусиків (антени) та ротові придатки. Антени — це багаточленикові придатки, що виконують функції органів дотику й нюху; у представників різних рядів їхня будова різна (рис. 119).
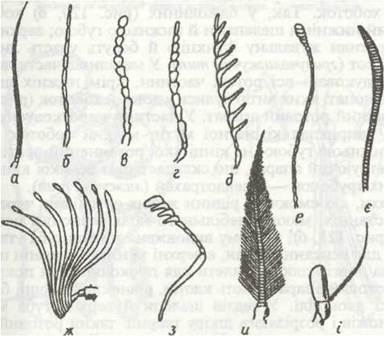
Рис. 119. Типи вусиків комах:
а - щетинкоподібний; б - ниткоподібний; в - чоткоподібний; г - пальчастий; д - гребінчастий; е - булавоподібний; є - веретеноподібний; ж - пластинчастий; з - колінчастий; й - пірчастий; і - щетинконосний
Ротові придатки, на відміну від покритощелепних, не занурені в ротову капсулу й складаються із нечленистої непарної верхньої губи (лабрум), пари нечленистих верхніх щелеп (мандибул), пари нижніх щелеп (максил) та непарної нижньої губи (лабіум), яка утворилася в результаті злиття
другої пари нижніх щелеп. Нижні щелепи та нижня губа двочленикові, несуть по парі органів дотику й смаку — щупиків. Залежно від характеру їжі розрізняють кілька типів ротових органів (рис. 120).
Гризучі (гризучожувальні) ротові органи (рис. 120, а) — найменш спеціалізовані, пристосовані для подрібнення твердої їжі. Верхня губа коротенька; верхні щелепи, як правило, мають жуйний край з більш-менш розвиненими зубцями. Зубці лівої щелепи входять у заглибини правої, і тому їхня будова не зовсім симетрична. Максили складаються з основного членика та стовпчика, на якому є пара нижньощелепних щупиків; він закінчується рухомо причленованими жуйними лопатями — зовнішньою та внутрішньою. Нижня губа має основний членик, або підборіддя, та дистальний членик, на якому є дві пари жуйних лопатей. До ротового апарата належить також м'ясистий виріст ротової порожнини — гіпофаринкс. Такі ротові органи характерні для тарганових, прямокрилих, термітів, бабок, твердокрилих та ін.
При переході до живлення рідкою їжею ротові органи значно змінюються залежно від того, як їжа розташована: чи відкрито (лижучий або сисний тип), чи схована під покривами (колючо- або ріжучосисні органи). У комах, які живляться відкрито розташованими рідинами, утворюється сисний хоботок. Так, у бджолиних (рис. 120, б) хоботок утворений нижніми щелепами й нижньою губою; верхні щелепи втратили жувальну функцію й беруть участь лише в побудові сот (гризучолижучий тип). У метеликів частково або зовсім редуковані всі ротові частини, крім нижніх щелеп, зовнішні лопаті яких витягнулись у довгий хоботок (рис. 121, а). Це сисний ротовий апарат. У частини коротковусих двокрилих, наприклад кімнатної мухи, м'який хоботок утворений нижньою губою, на кінці якої розвинений оригінальний фільтруючий апарат, що складається з великої кількості хітинових трубочок —псевдотрахей (лижучий тип).
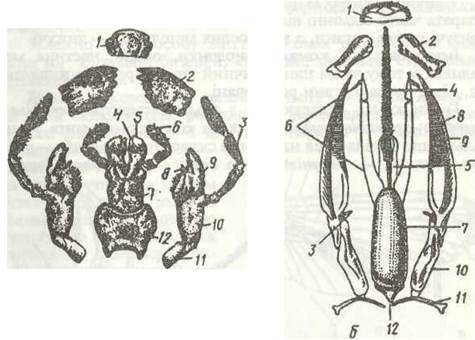
Рис. 120. Ротові апарати комах:
а - гризучий чорного таргана; б - гризучолижучий джмеля; 1 - верхня губа; 2 - верхня щелепа; 3 - нижньощелепний щупик; 4, 5 -внутрішня та зовнішня лопаті нижньої губи; 6 - нижньогубний щупик; 7 - підборіддя; 8, 9 -відповідно внутрішня та зовнішня лопаті нижньої шелепи; 10, 11 - відповідно стовпчик та основний членик нижньої шелепи; 12 — підпідборіддя
Комахи, які смокчуть рідини живих організмів через покриви останніх, мають здебільшого колючосисний ротовий апарат (рис. 121, б), в якому видовжена нижня губа утворює хоботок для всисання рідин, а верхні та нижні щелепи перетворені на довгі колючі стилети для проколювання покривів. Такий ротовий апарат мають клопи, рівнокрилі, воші, блохи, довговусі двокрилі. У ґедзів щелепи й верхня губа мають вигляд ножів і розрізають шкіру тварин: такий ротовий апарат зветься ріжучосисним. У мух-жигалок і це-це колючосисний апарат розвинувся з м'якого хоботка некровосисних предків, подібного до апарата кімнатної мухи. Він став твердим, а псевдотрахеї перетворилися на голку для проколювання шкіри.
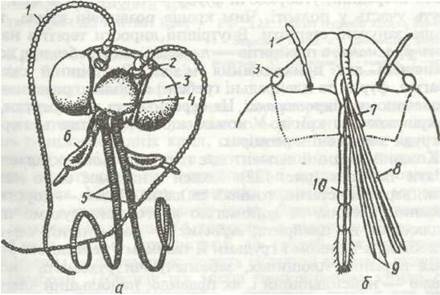
Рис. 121. Голови метелика із сисним ротовим апаратом (а) та клопа з колючосисним (б):
1 - антена; 2 - просте вічко; 3 - фасеткове око; 4 - лоб; 5 - хоботок; 6 - нижньогубний щупик; 7 - верхня губа; 8 - верхні щелепи;
9 - нижні шелепи; 10 - нижня губа
У личинок комах із повним перетворенням типи ротового апарата часто відмінні від таких у імаго: наприклад, у гусені гризучі ротові органи, а в дорослих метеликів — лижучі.
Імаго багатьох комах (одноденки, оводи, частина метеликів, у тому числі шовковичний шовкопряд) не живляться, і їхні ротові органи редуковані.
Груди комах складаються з трьох сегментів: передньо-, середню- та задньогрудей. Кутикула кожного сегмента — це кільце, що поділяється на чотири склерити: спинний —тер- гіт, грудний — етерніт та два бічні — плещипш. Тергіти зовні помітні краще, ніж етерніти, значна частина яких міс- титься всередині, утворюючи фурку — опору для м'язів, що беруть участь у польоті. Чим краще розвинені крила, тим більше занурені етерніти. Внутрішні вирости тергітів називають фрагмами, а плейритів —плейральними гребенями вони призначені для прикріплення м'язів. Внутрішній скелет (фрагми, фурки та плейральні гребені) особливо розвинений у так званому птеротораксі. Це середньо- та задньогруди, де прикріплюються крила. У комах, що добре літають, передньогруди зменшені в розмірах.
Кожний грудний сегмент має пару ніг. Нога складається з п'яти члеників (рис. 122). Кожен з них має свою назву: тазик, вертлюг, стегно, гомілка та лапка. Тазик — короткий основний членик, за допомогою якого нога рухомо прикріплюється до плейрита; вертлюг — найменший членик. Суглоби між тазиком і грудьми й тазиком і вертлюгом рухаються в різних площинах, забезпечуючи рухомість ноги. Стегно — найсильніший і, як правило, найбільший членик. Гомілка довга, але тонша, ніж стегно, часто озброєна шипами. Лапка в різних комах складається з одного-п'яти члеників і закінчується одним або двома кігтиками. У частини двокрилих на кінцевому членику, крім кігтиків, є присоски.
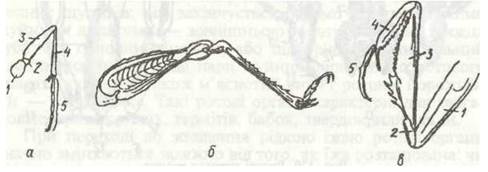
Рис. 122. Будова та типи ніг комах:
а - бігальна гуруна; б - стрибальна сарани; в - хапальна богомола; 1 - тазик; 2 - вертлюг; 3 - стегно; 4 - гомілка; 5 — лапка
За їх допомогою мухи повзають по вертикальних гладеньких поверхнях.
Залежно від способу життя в комах розвинулися різні типи ніг: ходильні, бігальні, копальні, хапальні, стрибальні, плавальні тощо.
Більшість комах має органи польоту — крила. Крила це бічні складки тіла, розташовані на середньо- та задньогрудях. Як звичайно, їх дві пари: передні та задні. Крило складається з двох стінок верхньої та нижньої. Кожна стінка утворена шаром гіподерми, зовні вкритої більш-менш розвиненою кутикулою (рис. 123). Між стінками є вузенька щілина (частина міксоцеля), заповнена гемолімфою.
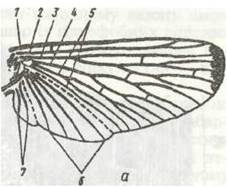
Рис. 123. Схема будови крила комахи:
а - загальний вигляд; б - поперечний розріз; 1 - костильна жилка, 2 - субкостальна; 3 - радіальна; 4 - медіальна; 5 - кубітальна; 6 — анальні, 7 - югальні жилки; 8 - кутикула; 9 - гіподерма; 10 - трахея
Крило (рис. 123, а) має систему хітинових трубочок-жилок. Кількість та взаємне розташування жилок відіграє велику роль у систематиці комах. Жилки виконують опорну функцію; в них також міститься гемолімфа, проходять трахеї та нерви до клітин крила.
Крила бувають різних типів. У частини комах (прямокрилі, твердокрилі, жорсткокрилі) передні крила перетворені на потовщені надкрила, що не беруть участі в польоті. Вони призначені для захисту ніжних задніх крил, складених під ними, при повзанні по землі, копанні в грунті тощо. У двокрилих задні крила перетворені на булавоподібні органи рівноваги — дзижнальця. У багатьох ґрунтових комах (робочі мурашки, терміти), а також у паразитів (воші, блохи) крила зникають, а в первиннобезкрилих (Apterygota) їх ніколи не було.
Крила рухомо прикріплені до грудей між тергітом та плейритом за допомогою досить складної системи склеритів та мембран. Поблизу від місця прикріплення крило спирається на виріст плейрита — стовпчик, який є для нього точкою опори і утворює важіль з коротким осьовим й довгим кінцевим плечима.
Черевце у найпримітивніших комах складається з 11 сегментів та тельсона, однак найчастіше їх буває вісім-дев'ять; у вищих груп (перетинчастокрилі, двокрилі) кількість їх може зменшуватися до чотирьох-п'яти. На VIII та IX сегментах розміщені зовнішні статеві придатки — геніталії, це копулятивний орган самців та яйцеклад у самиць.
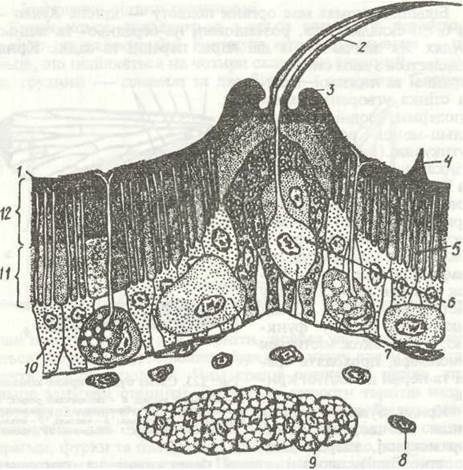
Рис. 124. Будова покривів комах:
1 - епікутакула; 2 - волосок; 3 - кільцевий валик біля основи волоска; 4 - шипик; 5 - перові канальці; 6 - клітина, що утворює волосок (трихогенна); 7 - базальна мембрана; 8 - гемоцит; 9 - жирове тіло; 10 - гіподерма; 11 - ендокутикула; 12 — екзокутикула
У зародковому стані на черевних сегментах є зачатки кінцівок, які надалі зникають чи видозмінюються у членисті придатки —церки (щетинкохвістки, таргани, одноденки) або нечленисті — грифельки (щетинкохвістки, таргани). Видозміненими кінцівками вважають також яйцеклад самиць, який у жалячих перетинчастокрилих перетворюється на жало. Геніталії самців складаються з різноманітних склеритів різної будови, походження яких неясне, однак парність частини з них указує на можливість їх виникнення від черевних кінцівок.
Покриви комах, як і решти членистоногих, мають три основні елементи: кутикулу, гіподерму і базальну мембрану (рис. 124). Кутикула утворює зовнішній скелет, що вкриває все тіло. Вона має поверхневий дуже тонкий шар — епікугикулу та внутрішній товстий — прокутикуду.
Епікутикула під світловим мікроскопом має вигляд напівпрозорої поверхневої лінії, проте вона неоднорідна й складається якнайменше з чотирьох шарів, що різняться за хімічним складом. Найглибший шар епікугикули — протеїновий, над ним послідовно один на одному лежать шари твердого кугикуліна, восковий шар із гідрофобних ліпідних сполук і тонкий цементний. Останній утворює лакове покриття кутикули. Характерна особливісь епікугикули — відсутність у її складі хітину.
Про кутикула утворена двома шарами: м'якою безбарвною ендокутикулою, що прилягає до гіподерми, й твердою, забарвленою пігментами екзокугикулою. Ендокутикула містить полімерні молекули хітину, зв'язаного з білками. Вони лежать шарами, складеними з найтонших пластинок, причому їхній напрямок у кожному з шарів змінюється (яку фанері), що робить ендокутикулу еластичною й водночас міцною. Екзокутикула не має пластинчастої будови. До неї, крім хітиново-білкових молекул, входять особливі дубильні речовини, що роблять її твердою (склеротизують), а також пігменти. Всю прокутикулу знизу доверху пронизують вертикальні перові канальці, всередині яких проходять тоненькі відростки гіподермальних клітин, їхній діаметр не перевищує 1 мкм, а загальна кількість досягає 10—15 тисяч на 1 мм2 покриву. Завдяки поровим канальцям відбуваються зв'язок гіподерми з кутикулою й утворення епікугикули під час линянь.
З усіх шарів кутикули ендокутикула найм'якша, гнучка, еластична, що пов'язано з її будовою. Це особливо важливо для місць зчленування кінцівок, крил, у міжсегментарних мембранах, де найбільш розвинений саме цей шар кутикули. Екзокутикула, навпаки, тверда, що залежить від ступеня задубленості — склеротизацїї хітиново-білкових комплексів фенольними сполуками. Цей шар краще розвинений там, де необхідна найбільша міцність: на тергітах та етернітах сегментів, у над крилах жуків, мандибулах, члениках кінцівок. На міжсегментарних мембранах екзокутикула відсутня. Вони цілком складаються з ендокутикули, вкритої зовні егакугикулою.
Епікутикула не дає випаровуватися воді з організму, утворюючи гідрофобний восковий шар. Ця її властивість особливо важлива для наземних комах, здатних жити на відкритому повітрі під сонячними променями, не втрачаючи воду свого організму. У комах та їхніх личинок, які живуть у воді, ґрунті, гниючій деревині, гноївці, епікутикули або зовсім немає, або вона вкриває лише певні ділянки тіла, наприклад головну капсулу.
Гіподерма складається з одного шару призматичних клітин, між якими трапляються залозисті клітини, що утворюють одноклітинні або багатоклітинні шкірні залози, а також спеціалізовані клітини, які утворюють волоски та сенсили. Зовнішня поверхня клітин гіподерми, що прилягає до кутикули, вкрита помітними лите під електронним мікроскопом мікроворсинками, які проникають у порозі канальці кутикули. Від порожнини тіла гіподерму відділяє базальна мембрана, яка не має клітинної будови. Найважливішою функцією гіподерми є секреторна, бо гіподерма виділяє речовини, з яких утворюється нова кутикула, причому багаторазово, під час линянь.
Кутикула комахи, яка щойно перелиняла, м'яка й безбарвна. У результаті склеротизації кутикула твердне; паралельно з цим процесом відбувається меланізація — синтез в екзокутикулі пігментів.
У комах розрізняють структурне й пігментне забарвлення. Структурне забарвлення пов'язане з деякими специфічними особливостями поверхневої структури кутикули мікроскопічними реберцями, пластинками, лусочками тощо, які створюють ефекти інтерференції, дифракції та розсіювання світла. Це металево блискучі та переливчасті забарвлення деяких жуків і метеликів, особливо тропічних. Пігментне забарвлення зумовлене пігментами, які найчастіше містяться в екзокутикулі, рідше — в клітинах гіподерми чи жирового тіла. В багатьох комах спостерігається поєднання структурного та пігментного забарвлення.
Найбільш поширеними пігментами комах є меланіни, що відкладаються в екзокутикулі й дають темно-коричневе, коричнево-червоне або чорне забарвлення. Часто в кутикулі комах трапляються каротиноїди, які створюють жовте, жовтогаряче, червоне забарвлення; флавоноїди жовтого кольору; білі, жовті та червоні птерини; жовті, коричневі або червоні омохроми. Антрахінони нагромаджуються в жировому тілі та гемолімфі комах ряду рівнокрилих (Homoptera), створюючи карміново-червоне забарвлення, що просвічує крізь покриви. До того як були синтезовані дешеві штучні барвники, червоний пігмент кармін добували з жирового тіла кошенілі Dactylopius coccus.
Покриви комах мають різноманітні придатки. Скульптурні придатки (шипики, виступи, боріздки, ямки тощо) утворюються кутикулою без участі гіподерми. Структурні придатки (волоски, щетинки, лусочки крил метеликів тощо) виникають одночасно як із кутикули, так і з гіподерми. Це або чутливі придатки, пов'язані з нервовими клітинами, або термоізоляційні утвори, що створюють густий покрив, подібний до волосяного (джмелі, деякі метелики та ін.).
Порожнина тіла — міксоцель комах — поділена двома поздовжніми горизонтальними перетинками —діафрагмами на три відділи (синуси). Верхня діафрагма відділяє верхній, або перикардіальний синус, в якому розташована спинна кровоносна судина (рис. 125). Нижня діафрагма відділяє нижній, або перинейральний синус, де міститься черевний нервовий ланцюжок. Середній синус лежить між діафрагмами; він називається вісцеральним; у ньому містяться травна, видільна й статева системи, а також більша частина жирового тіла. Порожнина тіла заповнена гемолімфою.
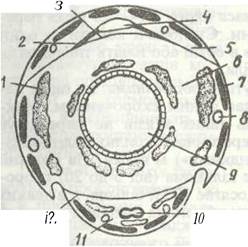
Рис. 125. Схема розташування серця та діафрагм на поперечному зрізі через тіло комахи:
1 - жирове тіло; 2 - крилоподібні м'язи; 3 - спинна діафрагма; 4 - серце; 5 - перикардіальний синус; 6 - периаісцеральний синус; 7-поздовжній м'яз; 8 -трахея; 9 - кишечних; 10 - нервовий ланцюжок; 11 - перинейральний синус; 12 -черевна діафрагма
Як скелетні м'язи, так і м'язи стінок внутрішніх органів (вісцеральні) комах виключно поперечносмугасті. Внаслідок високого ступеня розвитку органів руху та первинної переробки їжі м’язова система комах дуже диференційована. Наприклад, у гусені метеликів нараховується близько 2 тис. різних м'язів. Кожен м'яз за допомогою спеціальних волоконець — тонофібрил — кріпиться до двох різних склеритів, один з яких більш-менш фіксований, другий — · рухомий (рис. 126). Отже, скорочення м'язу спричиняє зміщення склеритів один відносно іншого.
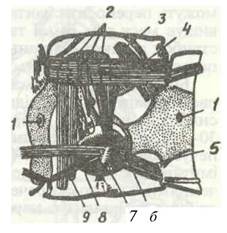
Рис. 126. Схема частини мускулатури задньогрудного сегмента комах:
1 - дихальце; 2 - дорзоветральні м'язи; 3 - косий спинний м'яз; 4 - поздовжній спинний м'яз; 5 — 8 - субкоксальні м'язи; 9 - поздовжній черевний м'яз
Скелетні м'язи утворюють головну, грудну та черевну групи. М'язи голови обслуговують ротові придатки, вусики й забезпечують рухомість шиї. М'язи грудей рухають ноги, а також крила. У черевці є поздовжні, бічні та поперечні м'язи, які надають йому рухливості; останні беруть участь в утворенні діафрагм, необхідних для роботи системи кровообігу.
Відносна сила м'язів комах дуже велика. Так, комахи можуть переносити вантаж, маса якого в 14—25 разів перевищує масу тіла самої тварини. Стрибаючі комахи за один стрибок долають відстань, що в сотні або навіть тисячі разів перевищує довжину їхнього тіла.
Більшість м'язів комах звуться синхронними: на один нервовий імпульс м'яз відповідає одним скороченням. Максимальна кількість скорочень таких м'язів не перевищує 30—40 за 1 с. У двокрилих і перетинчастокрилих політ забезпечується асинхронними («швидкими») м'язами. На кожний імпульс такий м'яз відповідає кількома (від 5 до 20) скороченнями, і число скорочень досягає 100 й більше, а в деяких дрібних двокрилих та перетинчастокрилих — навіть тисячі за секунду.
Типи живлення комах надзвичайно різноманітні: серед них є фітофаги та зоофаги, в тому числі хижаки, паразити й кровососи, сапрофаги, некрофаги (живляться трупами), копрофаги (живляться гноєм); є види, що живляться роговими утворами шкіри хребетних (шерсть, пір'я тощо), мертвою деревиною, воском тощо. Є також поліфаги — види, що вживають як рослинну, так і тваринну їжу. Способи живлення комах також дуже різні: одні поїдають тверді речовини, інші поглинають рідини, треті є фільтраторами. Цим зумовлюється різноманітність ротових органів. Сам кишковий тракт комах, хоч і модифікується залежно від способу живлення та складу їжі, але значно менше, ніж ротові органи.
Як і в решти членистоногих, травна система комах складається з трьох відділів: передньої, середньої та задньої кишок (рис. 127). Стінки всіх відділів кишечника утворені одношаровим епітелієм, зовні вкритим поздовжніми та кільцевими м'язовими волокнами, скорочення яких забезпечує рух їжі в кишечнику. Епітеліальні клітини передньої та задньої кишок на вільній поверхні вкриті кутикулярним шаром — інтимою.
Передня кишка складається з ротової порожнини, глотки, стравохода, вола та м'язового шлунка. У ротову порожнину відкриваються слинні залози, пов’язані з ротовими кінцівками. У комах бувають мандибулярні, максилярні та лабіальні (нижньогубні) залози. Найчастіше як слинні функціонують лабіальні залози, хоча в гусені метеликів слива утворюється з мандибулярних залозах, а лабіальні продукують шовк і не беруть участі в травленні. Слина змочує їжу та піддає її початковій дії ферментів, які розщеплюють полісахариди (крохмаль, глікоген). У попелиць слина містить ферменти, здатні перетравлювати целюлозні оболонки рослинних клітин; у кровосисних комах — антикоагулюючі речовини, що запобігають зсіданню крові. Слина комах-галоутворювачів містить різні специфічні амінокислоти та гормон росту рослин, які спричиняють утворення розростань рослинних тканин — галів.
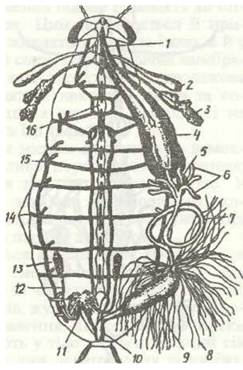
Рис. 127. Внутрішня будова самця чорного таргана при розтині зі спинної сторони:
1 - стравохід; 2 - резервуар слинної залози; 3 - слинна залоза; 4 -воло; 5 -жувальний шлунок; 6 - голоричні відростки середньої кишки; 7 - середня кишка; 8 - мальпігієві судини; 9 -товста кишка; 10 - пряма кишка; 11 - придаткові залози; 12 - сім'япровід; 13 - сім’яник; 14 -дихальця; 15- трахея; 16 - черевний нервовий ланцюжок
Глотка й стравохід забезпечують проковтування їжі та її проходження до вола.
Воло є місцем нагромадження їжі та початкового її перетравлювання під дією ферментів слини та травного соку, що потрапляє сюди із середньої кишки. В імаго двокрилих і лускокрилих замість вола є сліпий мішкоподібний виріст стравоходу — харчовий резервуар, у якому деякий час зберігається рідка їжа. Наприклад, у кровосисних двокрилих у ньому міститься випита вода або соки рослин. Кров сюди не потрапляє; вона проходить зі стравоходу прямо в середню кишку.
Жувальний шлунок має потужні м'язи; він вистелений усередині товстою кутикулою з гострими зубцями або товстими щетинками. У жувальному шлунку перетирається тверда їжа (прямокрилі, таргани, жуки) або фільтрується рідина (бджоли).
Середня кишка відділяється від передньої кардіальним клапаном — згорткою, що звисає в порожнину кишечника. Найпростіша вона у вигляді прямої трубки, не поліченої на відділи. У частини комах (мухи тощо) вона видовжена й звивиста, у деяких, навпаки, укорочена й розширена в мішкоподібний утвір (личинки перетинчастокрилих); у клопів вона має особливо складну будову й чітко поділяється на кілька відділів. У багатьох комах середня кишка утворює пілорити придатки — довгі або короткі пальцеподібні вирости (рис. 127, 128, а). Вони призначені для збільшення всисної поверхні кишечника й у деяких комах становлять притулок для симбіотичних мікроорганізмів.
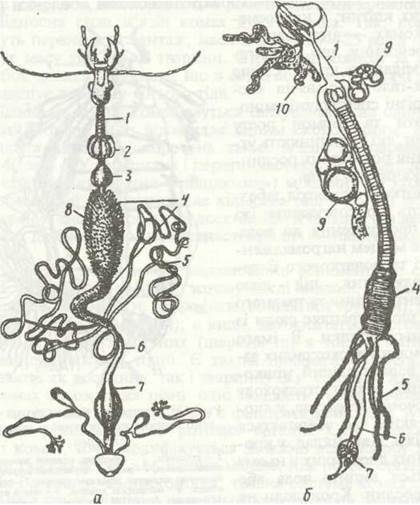
Рис. 128. Травна система жука туруна (а) та малярійного комара (б):
1 - стравохід; 2 - воло; 3 - жувальний шлунок; 4 - середня кишка; 5 - судини; 6 - задня кишка; 7 - пряма кишка; 8 -пілоричні придатки; 9 - харчові резервуари; 10 - слинні залози
У багатьох комах їжа, що надходить до середньої кишки, огортається тоненькою прозорою оболонкою —перитрофічною мембраною, що секретуєіься клітинами епітелія середньої кишки. Мембрана складається з білків і хітину й захищає стінку середньої кишки від механічних ушкоджень твердими частинками їжі. Крім того, вона має вибіркову проникність І регулює надходження продуктів травлення до клітин кишкового епітелію. Перитрофічна мембрана відіграє важливу роль у травленні: вона пропускає воду, мінеральні солі та продукти травлення, але затримує крупніші молекули білків, полісахаридів, ліпідів. Тому ферменти, що є всередині, не виходять за мембрану, чим досягається їх висока концентрація, а продукти травлення вільно підходять до стінок кишки, де й усмоктуються. Цим пояснюється її присутність не тільки в комах, які живляться грубою їжею, а й у тих, що ссуть кров чи рослинні соки. Перитрофічна мембрана відсутня в хижих жуків, для яких характерне позакишкове травлення, в комах, що живляться нектаром квітів та солодкими виділеннями попелиць, а також у комах, які на імагінальній фазі не приймають їжі (афатів).
Середня кишка є основним місцем травлення та всмоктування. У комах відбувається лише порожнинне травлення; внутрішньоклітинне травлення для них не характерне. У кишечнику їжа переміщується завдяки хвилеподібним скороченням м'язів його стінок: вони можуть спрямовувати їжу назад, вперед і перемішувати в певних ділянках.
У багатьох комах відбувається позакишкове травлення за допомогою слини. Так, тарган змочує свою їжу слиною, під дією якої вона розм'якшується та частково перетравлюється. Для деяких комах (хижих турунів, жуків-плавунців) характерне повністю позакишкове травлення. Ці комахи не тільки виливають слину, а й відригують у тіло здобичі травний сік середньої кишки і всмоктують уже перетравлену рідку їжу. Личинки мух, що живуть у трупах та гної, через анальний отвір виділяють травний сік із ферментами, які не тільки перетравлюють субстрат, а й убивають та лізують гнильні бактерії й гриби. З цим пов'язаний відкритий ще під час Кримської війни, в середині XIX ст., відомим російським хірургом М. Пироговим спосіб лікування гнійних ран. Стерильних (тобто одержаних у лабораторії й вільних від мікроорганізмів) личинок мух певних видів висаджували на гнійні рани. Вони виїдали змертвілі тканини і знищували всі мікроорганізми, не ушкоджуючи живих тканин, чим сприяли загоєнню ран.
Задня кишка відділена від середньої пілоричним клапаном і в більшості комах складається з тонкої, товстої та прямої (рекгум) кипток. На межі між середньою та задньою кишками в кишечник відкриваються тоненькі трубочки — мальпігієві судини, які виконують видільну функцію.
Задня кишка, як звичайно, не бере участі в травленні й не має ферментів, її функції пов'язані з формуванням екскрементів, водним обміном, виділенням та осморегуляцією.
Для багатьох комах їжа становить єдине джерело вологи, при цьому важливим є затримання її в організмі. У задній кишці відсмоктується вода з кишечника в гемолімфу (реабсорбція). У стінках прямої кишки є вирости — ректальні сосочки, які активно поглинають воду з порожнини кишечника й переводять її в гемолімфу. Крім води, ректальні сосочки вилучають із вмісту задньої кишки мінеральні іони (Na+, K+, Сl-), що забезпечує функцію осморегуляцїї. Задня кишка разом із мальпігієвими судинами функціонує як орган виділення (див. нижче).
У деяких комах травлення відбувається за допомогою симбіотичних мікроорганізмів. Наприклад, комахи, що живляться деревиною (терміти, деякі таргани, личинки пластинчастовусих жуків), не мають ферменту, який розщеплює клітковину. Його продукують симбіотичні найпростіші (джгутикові ряду Hypermastigida), бактерії та дріжджові гриби, що живуть у задній кишці. Вони розкладають целюлозу до оцтової кислоти, яка всмоктується ректальними сосочками. Отже, симбіотичне травлення відбувається в задній кишці. Симбіонти вола та середньої кишки різноманітні. Це бактерії, гриби, найпростіші, однак їх видовий склад і роль вивчено недостатньо. Відомо, що вони синтезують деякі вітаміни й амінокислоти. Наприклад, у південноамериканського кровосисного клопа Rhodnius prolixus фактично всі вітаміни, які в крові теплокровних майже відсутні, продукуються грибом-актиномцетом кишечника Nacardia rhodmi. Німфи цього клопа, позбавлені актиноміцетів, незважаючи на нормальне живлення кров'ю, не досягають фази імаго й гинуть.
Виділення в комах здійснюється кількома органами, які не створюють єдиної системи. Це мальпігієві судини, задня кишка, уратні клітини жирового тіла, перикардіальні клітини та деякі специфічні утвори окремих груп комах. Основними видільними органами є мальпігієві судини й задня кишка, що функціонують як єдине ціле. Мальпігієві судини було відкрито в XVII ст. італійським ученим Мальпігі. У типовому випадку це довгі тоненькі трубочки, які впадають у кишечник на межі середньої та задньої кишок (рис. 129, а). Протилежні сліпозамкнені кінці їх вільно плавають у гемолімфі, їхня кількість коливається в різних комах від 2 до 200; у попелиць вони зникли внаслідок редукції.
Деталі будови мальпігієвих судин різняться в окремих групах комах: у клопів вони зрощені кінцями попарно, утворюючи петлі; у комах, яким особливо необхідне заощадження води, наприклад у гусені метеликів, котра одержує воду лише з їжі, кінці мальпігієвих судин приростають до задньої кишки (рис. 129, б), чим забезпечується додаткове всмоктування води з останньої.
Стінки судин утворені одношаровим епітелієм, зовні вкритим базальною мембраною та м'язовими волокнами. Скорочення м'язів спричиняють рух судин у гемолімфі, а також перистальтичні та антиперистальтичні рухи, необхідні для перемішування екскретів і проштовхування їх у кишечник.
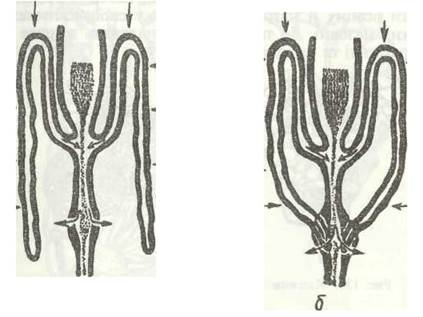
Рис. 129. Основні типи мальпігієвих судин комах (стрілками показано рух екскретів, води та мінеральних іонів):
а - прямокрилих; б - жуків
Мальпігієві судини всмоктують гемолімфу з продуктами обміну. Це водний розчин вуглеводів, амінокислот і солей сечової кислоти. Він функціонально відповідає первинній сечі хребетних. З уратів, розчинних солей основного продукту азотистого обміну комах —сечової кислоти —у мальпігієвих судинах утворюється малорозчинна сечова кислота. У задній кишці ректальні сосочки вилучають із цієї рідини й повертають до гемолімфи більшу частину води, поживні речовини та іони неорганічних сполук. Зневоднені кристали сечової кислоти разом із фекаліями виводяться назовні через анальний отвір.
Видалення з організму продуктів азотистого обміну у вигляді нерозчинної у воді сечової кислоти — важливе пристосування комах до життя в умовах дефіциту вологи. У більшості водяних безхребетних основним екскретом є аміак, добре розчинний у воді, але дуже токсичний: навіть у низьких концентраціях він може спричиняти тяжкі отруєння. Тому такий спосіб виділення доступний лише водяним тваринам, яким не доводиться заощаджувати воду. Для наземних комах, особливо таких, що живуть на відкритому повітрі (метелики та їхні личинки, більшість дорослих жуків, перетинчастокрилих, двокрилих тощо), важливим є збереження вологи, тому вони виділяють майже сухі екскрети, які на 90 % складаються з сечової кислоти. Показово, що комахи або їхні личинки, які перейшли до життя у воді або іншому рідкому середовищі (личинки падальних мух), виділяють аміак.
Крім мальпігієвих судин і задньої кишки, видільну функцію виконують також органи, що вилучають із гемолімфи продукти обміну й нагромаджують їх у своїх клітинах без виведення назовні. До таких органів належать жирове тіло, перикардіальні та гіподермальні клітини.
У жировому тілі, крім клітин-трофоцитів, які запасають поживні речовини, трапляються також уратні клітини, що нагромаджують сечову кислоту (рис. 130). Нагромаджувальна функція цих клітин має важливе значення в ті періоди розвитку комах, коли виведення екскретів назовні неможливе. Наприклад, в ендопаразитичних личинок екскреторні продукти відкладаються в жировому тілі, оскільки виведення їх в організм хазяїна може спричинити отруєння останнього й загибель самого паразита. Екскрети видаляються назовні лише після вильоту імаго. У лялечок вищих двокрилих задня кишка та мальпігієві судини руйнуються й не функціонують, а сечова кислота акумулюється в уратних клітинах жирового тіла.
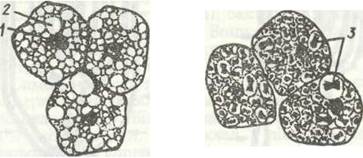
Рис. 130. Клітини жирового тіла личинки комара Aedes aegypti:
а - трофоцити; б - уратні клітини; 1 - ядро; 2 - жирова вакуоля; 3 - кристали сечової кислоти
Перикардіальні клітини, що оточують спинну кровоносну судину, здатні поглинати великі білкові молекули та різні колоїдні частинки, які потрапляють у гемолімфу. Клітини гіподерми також можна вважати органами нагромаджувальної ексіфеції, оскільки азотисті продукти обміну речовин використовуються тут для синтезу хітину кутикули та ΤΓ- менгів — меланінів, омохромів, птеринів.
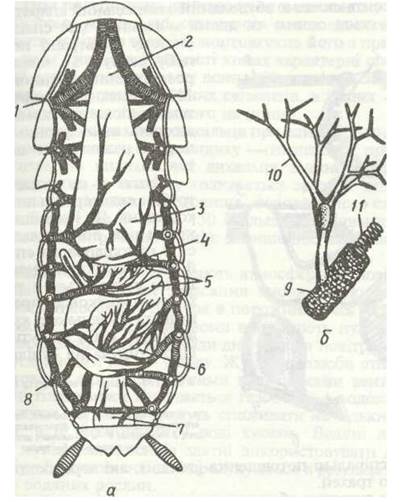
Рис. 131. Трахейна система чорного таргана:
а - загальний вигляд зі спинної сторони; б - закінчення трахей з трахеолами; 1 - грудні стигми; 2 - стравохід; 3 - воло; 4 - жувальний шлунок; 5 - пілоричні відростки; 6 - середня кишка; 7 - пряма кишка; 8 - черевні стигми; 9 -трахея; 10 - трахеоли; 11 — тенідій
У деяких комах видільну функцію виконують ще лабіальні залози: у щетинкохвісток (підклас Apterygota) вони виділяють барвники, у метеликів родини Satumiidae — розчин бікарбонату калію. Додаткові залози статевих органів самців тарганів також виводять значну кількість сечової кислоти. Інколи сечова кислота нагромаджується в кутикулі, зумовлюючи її біле забарвлення, наприклад, у крилевих лусочках метеликів — біланів (родина Pieridae).
Дихальна система комах представлена трахеями (рис. 131). Трахеї відкриваються назовні кількома парами дихалець — стигм. Дві пари стигм розташовані відповідно на середньо- та задньогрудях; на перших восьми сегментах черевця лежить по парі стигм, однак кількість їх може зменшуватися. Стигми мають досить складно збудований замикальний апарат, який обслуговується одним чи двома м'язами й має спеціальну
систему фільтрації повітря, побудовану з численних розгалужених щетинок (рис. 132). Описана трахейна система типова для комах, які дихають атмосферним киснем, і називається відкритою.
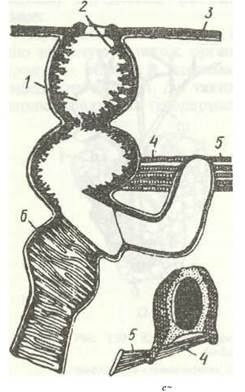
Рис. 132. Один із варіантів будови дихальця:
а - поздовжній, б - поперечний зрізи; 1 - атріальна порожнина; 2 - фільтрувальний апарат; 3 - кутикула; 4 - м'яз-замикач дихальця; 5 - м'яз-відкривач; 6 -трахея
Трахеї — це розгалужені трубки ектодермального походження. Як і зовнішні покриви тіла, вони складаються з одношарового епітелію й вистелені кутикулою. Остання складається з товстої хітиново-білкової прокугикули та тоненького кутикулинового шару епікутикули. Прокутикула не суцільна, а утворює спіральні потовщення (тенідії), які перешкоджають злипанню трахей.
Кожне дихальце обслуговує три поперечні трахеї, з'єднані між собою трьома парами поздовжніх трахей. Від цих основних стовбурів відходять розгалуження до всіх органів. Вони закінчуються тоненькими трубочками діаметром 1— 2 мкм — трахеолами. Кінці трахеол або лежать на поверхні окремих клітин, або входять усередину їх. Кисень із трахеол безпосередньо дифундує в клітини, а вуглекислий газ — із тканин у трахеоли.
У комах, що добре літають, на поздовжніх трахейних стовбурах утворюються розширення — повітряні мішки. Вони не мають тенідіїв і можуть змінювати об'єм. Повітряні мішки беруть участь у вентиляції крилевої мускулатури під час польоту й виконують аеростатичну функцію, сприяючи зменшенню питомої маси тіла.
Кисень із повітря, яке заповнює трахейну систему, транспортується до окремих клітин тіла шляхом дифузії. В клітинах він одразу ж споживається, тому в трахейній системі виникають дифузійні струми кисню, спрямовані всередину тіла. Крім того, в багатьох комах є додаткова вентиляція трахей. При польоті вона здійснюється в основному за рахунок крилових м'язів, які синхронно з коливаннями крил нагнітають повітря в трахеї й виштовхують його з повітряних мішків до м'язів. Для більшості комах характерні спеціальні дихальні рухи: в одних комах вони здійснюються за рахунок ритмічних розширень черевних сегментів, в інших — телескопічного насування їх одного на інший.
У більшості комах одні дихальця при вдиху відчиняються, інші — зачиняються, а при видиху — навпаки. У проміжках між вдихом та видихом усі дихальця зачинені. Кількість дихальних рухів за хвилину коливається залежно від температури навколишнього середовища, фізіологічного стану комахи та її виду від 5—6 до 150 і більше. Закривання стигм між дихальними рухами сприяє зменшенню випаровування води.
У водяних комах, які дихають атмосферним повітрям, є спеціальні пристрої для запасання кисню. Жуки-плавунці набирають атмосферне повітря в порожнину між черевцем і надкрилами. З-під надкрил вони випускають пухирець повітря, через який кисень із води дифундує в повітряну камеру, а вуглекислий газ — у воду. Жуки-водолюби створюють запас повітря між гідрофобними волосинками вентральної поверхні тіла, де теж відбувається газообмін з водою. Отже, водяні жуки під водою можуть споживати не тільки атмосферний, а й розчинений у воді кисень. Водяні личинки деяких жуків-довгоносиків здатні використовувати для дихання бульбашки кисню, які продукуються в результаті фотосинтезу водяних рослин.
У багатьох ендопаразитичних личинок трахейна система частково чи зовсім редукована, й дихання здійснюється через покриви. Деякі з них під'єднують свою трахейну систему до трахей комахи-хазяїна, інші проривають його покриви, виставляючи дихальця назовні.
У багатьох водяних або ендопаразитичних видів трахеї назовні не відкриваються (замкнена трахейна система); вони дихають киснем, розчиненим у воді чи в рідинах тіла хазяїна. У цих комах дихання відбувається або через усю поверхню тіла, або за допомогою спеціальних органів. У водяних личинок або німф дихальця не відкриваються назовні, а продовжуються в трахеї, розгалужені в тонкостінних плівчастих або гілчастих виростах — трахейних зябрах (рис. 133). Трахеї наповнені повітрям, і газообмін здійснюється з водою через зяброву поверхню. У личинок одноденок, жуків-вертячок, волохокрильців тощо такі зябра метамерно розташовані на черевці; у німф різнокрилих бабок зябра містяться всередині задньої кишки. Німфа періодично вбирає та викидає воду з кишки за допомогою ректальних м'язів; водночас із вентиляцією відбувається реактивний рух тварини вперед при викиданні з анального отвору струменя води.
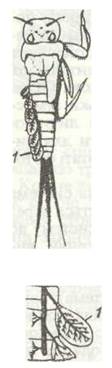
Рис. 133. Трахейні зябра личинки одноденки (а) та схема їх трахеації (б): 1 - зябра; 2 - трахея
Безпосередній транспорт газів через трахеї до тканин і клітин енергетично значно вигідніший, ніж багатоетапна система дихання хребетних (органи дихання — кров — міжклітинна рідина — тканини), проте ефективна лише при малих розмірах тіла, а в разі збільшення біомаси м'язи не здатні накачати достатню кількість повітря в клітини. Саме завдяки безпосередній доставці кисню до клітин можуть функціонувати асинхронні крилові м'язи. У м'язах хребетних тварин дефіцит кисню під час посиленої роботи призводить до їх утоми.
Кровоносна система комах дуже редукована через майже повну втрату гемолімфою функції транспорту газів. Від неї залишається спинна судина, розташована в перикардіальному синусі й підвішена за допомогою сполучнотканинних тяжів до спинної стінки тіла. Задня її частина — серце, передня — аорта (рис. 134). Серце складається з ряду послідовних камер і розташоване в черевній тагмі. Кожна камера серця має пару бічних отворів — остій з клапанами. Через них гемолімфа потрапляє з перикардія всередину серця. Клапани перешкоджають її зворотному руху. Камери сполучені між собою отворами, у частини комах — із клапанами, що не дають змоги крові рухатись назад. Задній кінець серця замкнений, передній — подовжений у трубчасту аорту, яка відкривається в міксоцель поблизу голови. До верхньої діафрагми та нижньої сторони кожної камери прикріплюється пара крилоподібних м'язів.
Камери серця почергово розширюються (діастола), і гемолімфа через остії з перикардія надходить у серце, а потім звужуються (систола), і гемолімфа тече вперед. Пульсація серця спричиняється еластичністю його стінок, а також роботою крилоподібних та інших м'язів. З аорти гемолімфа потрапляє в порожнину голови, де утворюється зона підвищеного тиску. Відповідно в задній частині тиск гемолімфи менший, тому вона по середньому й нижньому синусах тече назад, а по спинній судині знову повертається вперед. Частота серцевих скорочень залежить від виду комахи, її фізіологічного стану, фази розвитку та впливів факторів зовнішнього середовища й коливається від 10 до 150 скорочень за хвилину.
Біля основи антен, ніг, крил є місцеві пульсуючі органи, які нагнітають у них гемолімфу. Найчастіше це м'язові ампули, скоротливі перетинки, які пульсують незалежно від ритму серця. Жилки крил, поздовжні перетинки в кінцівках (септи) утворюють упорядковану систему руху гемолімфи. Дихальні рухи також сприяють її циркуляції.
Гемолімфа комах складається з рідкої міжклітинної речовини — плазми та клітин —гемоцитів, які або плавають у плазмі, або нерухомо осідають на поверхні внутрішніх органів. У більшості комах в 1 мм3 гемолімфи міститься від 10 000 до 100 000 клітин, а їхній загальний об’єм досягає 10 % об'єму гемолімфи.
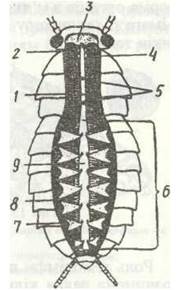
Рис. 134. Схема кровоносної системи таргана:
1 - аорта; 2 - кардіальні тіла; 3 - мозок; 4 - прилеглі тіла; 5 - розгалуження крилових кровоносних судин; б - серце; 7 - діафрагма; 8 - крилоподібні м'язи; 9 -серцеві камери
Плазма гемолімфи — це водний розчин неорганічних та органічних речовин. У ній є неорганічні іони та амінокислоти, які беруть участь у підтримці водно-сольового балансу й осморегуляції. Плазма гемолімфи містить також вуглеводи, органічні кислоти, гліцерин, ліпіди, пептиди, білки та пігменти.
Гемоцшпи — це клітини мезодермального походження. Усі вони безбарвні й мають ядра. Розрізняють кілька типів гемоцитів (рис. 135): одні з них можуть утворювати псевдоподії й здійснювати фагоцитоз, інші — нагромаджують поживні речовини, наприклад глікоген, і транспортують їх у тканини. У різних ділянках тіла, переважно в жировому тілі, є скупчення недиференційованих клітин, які перетворюються на гемоцити й потрапляють у плазму.
Гемолімфа утворює рідке внутрішнє середовище організму. Вона виконує деякі важливі функції. Перша — це транспорт поживних речовин, гормонів та інших біологічно активних речовин, а також продуктів обміну до відповідних органів, тканин і клітин. Друга важлива функція — захист організму від інфекційних та інвазійних захворювань. Третя — підтримка сталості хіміко-фізичних властивостей внутрішнього середовища організму. Важлива й механічна функція: внаслідок гідростатичного тиску гемолімфи змінюється форма органів з м'якою кутикулою —розправляються крила в імаго після виходу з лялечки, розкручується хоботок метеликів тощо.
Рис. 135. Деякі типи гемоцитів комах:
а - недиференційованз клітина; б - фагоцитуюча клітина; в, г —клітини з білково—тікогенними та жировими резервами
Роль гемолімфи в транспорті газів незначна, однак у ній розчинена деяка кількість кисню, необхідного для дихання гемоцитів. У личинок комарів родини Chironomidae в гемолімфі є розчинений гемоглобін, але тут він виконує функцію запасання, а не транспортування кисню. Завдяки наявності зв'язаного гемоглобіном кисню хірономіди можуть надовго зариватися в мул, використовуючи для дихання кисень, який відщеплюється від гемоглобіну.
Гемолімфа має здатність до зсідання. При ушкодженні покривів вона витікає назовні й утворює згусток з гемоцитів та плазми, який закриває рану.
Частина гемоцитів здатна до фагоцитозу. Вони поїдають і перетравлюють шкідливі мікроорганізми, що обумовлює стійкість комах проти хвороботворних мікроорганізмів.
Неспецифічні імунні реакції забезпечують фагоцитуючі гемоцити, а також плазма, де є комплекс ферментів та антибіотиків із широким спектром дії на різні мікроорганізми, наприклад фермент лізоцим, який руйнує оболонки бактеріальних клітин. Крім того, гемоцити утворюють капсули навколо багатоклітинних паразитів (нематоди, личинки їздців тощо), що призводить до загибелі останніх. Специфічний імунітет для комах не характерний, антитіла в гемолімфі не утворюються.
Гемолімфа багатьох комах отруйна й використовується для захисту від ворогів. У разі небезпеки вона виділяється назовні через суглоби кінцівок і вусиків. Це явище відомо для жуків-сонечок (Coccinellidae), листоїдів (Chrysomelidae). Жук шпанська мушка (Lytta vesicatoria) у гемолімфі має отруйну речовину кантаридин, яка спричиняє запалення шкіри й загальну інтоксикацію хребетних.
Тісно пов'язане з гемолімфою жирове тіло, яке разом з нею утворює внутрішнє середовище організму. Ця пухка тканина мезодермального походження складається з численних лопатей між внутрішніми органами (рис. 136). Клітини жирового тіла за будовою та походженням близькі до гемоцитів. Більшість клітин жирового тіла становлять трофоцити. У них нагромаджуються резервні поживні речовини —жири, білки та глікоген. Крім того, як уже згадувалося, в ньому є уратні клітини, в яких нагромаджуються кристали сечової кислоти (див. рис. 130).
Рис. 136. Зріз через лопать жирового тіла чорного таргана: 1 - жирові клітини; 2 - клітини із симбіотичними бактеріями
Основною функцією жирового тіла є нагромадження резервів поживних речовин на личинковій фазі розвитку та забезпечення ними організму на час метаморфозу, діапаузи (див. нижче), голодування, дозрівання статевих продуктів. При цьому жирове тіло не тільки пасивно нагромаджує поживні речовини, а й здійснює проміжний обмін речовин. У його клітинах відбуваються процеси біосинтезу та перетворення білків, жирів, вуглеводів. У самиць під час розмноження жирове тіло синтезує специфічні білки — вітелогеніни, необхідні для утворення жовтка в яйцях, що розвиваються. У багатьох комах у жировому тілі є особливі клітини — міцетоцити (рис. 136), у цитоплазмі яких живуть симбіотичні мікроорганізми — бактерії та гриби, що продукують деякі вітаміни та інші біологічно активні речовини, необхідні комахам. Міцетоцити згруповані у скупчення, що називаються міцетомами.
Деякі комахи, наприклад жуки-світляки (родина Lampyridae), мають органи свічення (фотогенні органи) — видозмінені ділянки жирового тіла (рис. 137). Вони залягають під прозорим кутикулярним покривом черевця імаго, інколи личинок. Свічення залежить від наявності в клітинах особливої речовини — люциферину. Під впливом нервового імпульсу люциферин у присутності ферменту люциферази та іонів Mg реагує з АТФ, перетворюючись на люциферин-аденілову кислоту, яка негайно окислюється киснем і виділяє світло. У дорослих комах взаємні світлові сигнали певної частоти призначені для знаходження особинами різної статі одне одного.
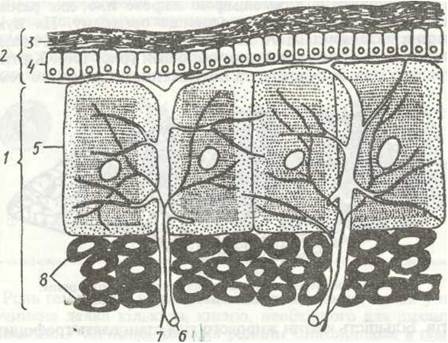
Рис. 137. Схема будови органа свічення світляка:
1 - жирове тіло; 2 - покриви; 3 - кутикула; 4 - гіподерма; 5 - фотоцит; 6 - трахея; 7 - нервове волокно; 8 - клітини, що відбивають світло
Центральна нервова система комах, як і інших членистеногих, складається з парного надглоткового ганглія, або головного мозку, навкологлоткових конектив та черевного нервового ланцюжка. Перший ганглій ланцюжка — підглотковий — лежить разом із надглотковим у голові, решта —в тулубі.
Надглотковий ганглій (рис. 138) складається з трьох злитих разом гангліїв: протоцеребрума, дейтоцеребрума та тритоцеребрума.
Протоцеребрум, або передній мозок, розвинений краще, ніж інші, й має найскладнішу будову. У ньому розрізняють кілька гангліозних центрів, серед яких найкраще розвинена пара стебельчастих, або грибоподібних, тіл —вищого асоціативного та координуючого центру нервової системи. Вони досягають найвищого розвитку в комах зі складними формами поведінки, особливо в перетинчастокрилих. Крім того, в протоцеребрумі міститься пара великих зорових часток, які іннервують складні очі.
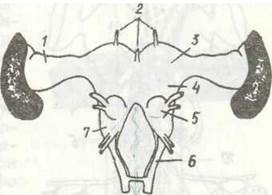
Рис. 138. Надглотковий ганглій богомола:
1 - зорові частки протоцеребрума; 2 - нерви дорзальних вічок; 3 - протоцеребрум; 4 - дейтоцеребрум; 5 - нюхові частки дейтоцеребрума; 6 - навкологлоткові конекгивн; 7 - тритоцеребрум
Дейтоцеребрум — середній мозок —містить парні нюхові центри, він іннервує антени.
Тритоцеребрум — задній мозок — іннервує верхню губу. З ним пов'язана вегетативна (симпатична) нервова система.
Підглотковий ганглій іннервує ротові органи та слинні залози.
Черевний нервовий ланцюжок у примітивніших комах (прямокрилі, таргани) складається з трьох грудних і восьми черевних гангліїв. В інших комах черевних гангліїв менше, що пов'язано з концентрацією нервової системи. Скорочення кількості гангліїв досягається в результаті об'єднання як черевних, так і грудних гангліїв, і у вищих груп комах приводить до злиття всіх гангліїв у два-три або навіть один великий ганглій, наприклад у вищих мух і жуків.
Крім центральної, у комах добре розвинена вегетативна нервова система. Вона складається з трьох відділів: стоматогастричного (ротошлункового), вентрального, або черевного, та каудального (хвостового).
Стоматогастричний відділ (рис. 139) складається з кількох самостійних гангліїв і нервів (фронтальний, потиличний, шлунковий ганглії, поворотний нерв), але має зв'язки з мозком. Стоматогастрична система іннервує серце й передню частину кишечника.
Вентральний, або черевний, відділ складається з непарного нерва, який тягнеться паралельно до черевного нервового ланцюжка вздовж усього тіла; його вищим центром є тритоцеребрум. Вентральний нерв іннервує дихальця, трахеї, жирове тіло; разом із гангліями центральної нервової системи він посилає нерви до м'язів, справляючи на них регулювальний вплив. Після його руйнування спостерігається швидке втомлення крилевих м'язів.
В останньому черевному ганглії непарний нерв розпадається на дві гілки, які іннервують задню кишку й статеві органи, що його частину називають каудальним відділом.
Зі стоматогастричним відділом тісно пов'язані мозкові залози —кардіальні й прилеглі тіла. Це невеличкі парні тільця, розташовані позаду мозку. За допомогою спеціальних нервів кардіальні та прилеглі тіла з'єднуються з мозком і потиличним ганглієм. Вони входять до складу ендокринної системи, куди належать також нейросекреторні клітини й проторакальні (передньогрудні) залози (рис. 139).
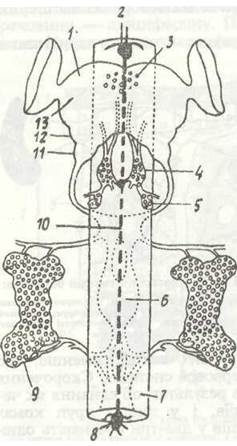
Рис. 139. Схема ендокринної та вегетативної (стоматогастричної) систем комах:
1 - надглотковий ганглій; 2 - фронтальний ганглій; 3 - нейросекреторні клітини; 4 - кардіальні тіла; 5 - прилеглі тіла; 6 - ганглій черевного нервового ланцюжка; 7 - кишечник; 8 - шлунковий ганглій; 9 - проторакальні залози; 10 - поворотний нерв; 11 - тритоцеребрум; 12 - дейтоцеребрум; 13 - протоцеребрум
Ендокринні органи синтезують, нагромаджують і виділяють у гемолімфу гормони, які регулюють усі фізіологічні процеси в організмі.
Усі ганглії центральної нервової системи мають у своєму складі нейросекреторні клітини. Найбільше їх у протоцеребрумі (рис. 139). Вони продукують мозковий, або активаційний, гормон, який активізує інші ендокринні органи, стимулюючи виділення ними гормонів. Нейросекреторні клітини підглоткового ганглія продукують гормон ембріональної діапаузи. Нейросекреторні клітини гангліїв черевного нервового ланцюжка виділяють бурсикон — гормон, який спричиняє затвердіння (склеротизацію) кутикули під час линяння.
Кардіальні тіла нагромаджують і виділяють нейрогормони, а також виробляють гормони, що регулюють вуглеводний і жировий обмін. Прилеглі тіла виробляють ювенільний гормон, який на фазі личинки сприяє розвитку личинкових
органів, але гальмує перетворення на імаго. У самиць дорослих комах цей гормон стимулює розвиток яєць.
Проторакальні залози — це пара залоз, що лежать у черевній частині передньогрудей по обидва боки передньо-грудного ганглія і зв'язані з ним нервами. Вони виділяють гормон линяння, або екдізон.
Органи чуття комах — найскладніші й найрізноманітніші, що пов'язане із загальним високим рівнем організації та складною поведінкою комах, яка вимагає точної інформації про навколишній світ. Комахи здатні сприймати різноманітні подразнення й мають такі рецептори: механорецептори (сприймають дотик, вібрацію та звукові хвилі); терморецептори (реагують на зміну температури); гігрорецептори (реагують на вологу); хеморецептори (сприймають хімічні стимули); фоторецептори (сприймають світлові подразнення). Є ще пропріоцептори, які сигналізують нервовій системі про положення, деформацію та зміщення окремих ділянок тіла.
Морфологічну й функціональну основу чутливості комах становлять нервово- чутливі одиниці — сенсили.
Вони або розкидані по різних частинах тіла, або зібрані у скупчення — органи чуття (очі, органи слуху тощо). Як і в інших членистоногих, сенсила комах складається з кутикулярної частини, однієї або кількох чутливих клітин та обслуговуючих клітин (рис. 140). Залежно від форми і розташування кутикулярних частин розрізняють трихощні, базиконічні, целоконічні, дзвоноподібні, плакоїцні та інші сенсили (рис. 141).
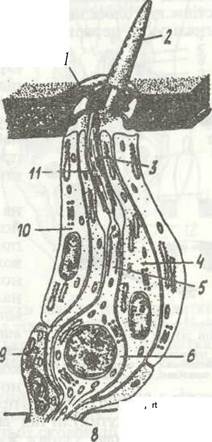
Рис. 140. Ультраструктурна організація трихоїдної механорецепторної сенсили:
1 - зчлековна мембрана; 2 - волосок; 3 - кутикулярна оболонка; 4 - трихогенна клітина; 5 - дендрит чутливої клітини; 6 - чутлива клітина; 7- базальна мембрана; 8 -аксон; 9 - гліальна клітина; 10 - тормогекна клітина; 11 - нерухомий джгутик
Рис. 141. Кутикулярні частини різних типів сенсил:
а - трихощної; б - базиконічної; в - стилоконічної; г - целоконічної; д - плакощноі; е - ампулоподібної
До механорецепторів належать дотикові рецептори, а також структури, що сприймають коливання субстрату, вітру або власного тіла комахи, його положення тощо. Найпростішими механорецепторами є трихоїдні сенсили. Вони розкидані по всьому тілу, але найбільше їх на тих частинах тіла й придатків, які найчастіше контактують із оточуючими предметами (антенах, ногах, яйцекладі тощо). Особливий різновид становлять трихоїдні сенсили, розташовані найчастіше на голові та крилах — вітрочутливі рецептори. Вони сигналізують нервовим центрам про початок, інтенсивність, тривалість і напрямок повітряних струмів, які обдувають тіло комахи під час польоту. У тарганів та цвіркунів такі сенсили містяться на церках і сигналізують про швидке наближення до них будь-якого предмета, що спричиняє реакцію втечі.
Механорецептори, які реагують на зміщення сегментів тіла та рух його придатків, належать до пропріоцепторів. Вони представлені волосковими пластинками, дзвоноподібними сенсилами, хордотональними органами та рецепторами розтягнення. Волоскові пластинки — це скупчення трихощних сенсил, які розміщені в місцях контакту сегментів тіла, члеників ноги, антен тощо (рис. 142). Дзвоноподібні сенсили слугують пропріоцепторами, які реагують на деформацію кутикули під час скорочень м'язів, особливо їх багато на крилах уздовж жилок, на ногах, яйцекладі, мандибулах.

Рис. 142. Волоскова пластинка в місці зчленування ноги з тілом тар
1 - плейрит тулубного сегмента; 2 - волоскова пластинка; 3 -тазик ноги
Хордотональні органи — це сукупність особливих механорецепторних сенсил (сколопіціїв), натягнутих між двома ділянками кутикули. За своєю будовою сколопщії відрізняються від інших сенсил (рис. 143). їхню основу складає нервова клітина, чутливий відросток якої (довгий нерухомий джгутик) оточений по всій довжині кутикулярним чохлом — штифтом, або сколопсом, що є продуктом виділення облямовуючої клітини. Дистальний кінець джгутика входить у канал шапочки, оточеної шапочковою клітиною. Остання прикріплюється до кутикули.
Хордотональні органи реагують на деформацій кутикули, спричинені рухами комахи. Натискання шапочки на дистальний кінець джгутика збуджує чутливу клітину. Хордотональні органи виконують функції пропріоцепторів і сигналізують нервовій системі про рухи тіла та його придатків. Вони містяться на різних частинах тіла — в ротовому апараті, антенах, грудях, ногах, крилах, черевці та його придатках. Деякі Хордотональні органи чутливі до вібрацій та сильних звуків.
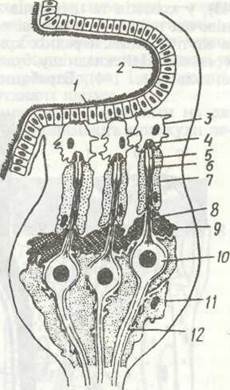
Рис. 143. Група з трьох сколопідіїв у тимпанальному органі сарани:
1 - тонка і жорстка та 2 - товста і м'яка ділянки тимпанальної мембрани; 3 - шапочкова клітина; 4 - шапочка; 5 - нерухомий джгутик; 6 - сколопс; 7 - облямовуюча клітина; 8 - дендрит; 9 - фіброзна клітина; 10 - чутлива клітина; 11 - гліальна клітина; 12 - аксон чутливої клітини
Особливим різновидом хордотональних органів є джонстонів орган, що міститься в другому членику антен майже всіх комах. Він складається з багатьох сколопідіїв, натягнутих між стінками другого членика антени та зчленовною мембраною, яка з'єднує його з третім члеником. Сколопідії реагують на найменші коливання антени при рухах тварини, коливаннях і струсах повітря або субстрату. У кровосисних комарів (родина Culicidae) джонстонів орган має кілька тисяч сколопідіїв і виконує функцію органа слуху, який сприймає звукові коливання високої частоти.
Слух розвинений не в усіх комах. Найчастіше слухові органи мають ті з них, які самі здатні створювати звуки. Спеціалізовані органи слуху звуться тимпанальними органами. Вони подібні до хордотональних, але відрізняються від останніх тим, що сколопідії в них прикріплюються до витонченої у вигляді барабанної перетинки ділянки кутикули й сприймають її коливання під дією звукових хвиль. У саранових вони лежать по боках першого сегмента черевця (рис.143), у коників та цвіркунів — на гомілках передніх ніг, у співочих цикад — в основі черевця, у денних метеликів на здутій основі передніх крил, у совок — між грудьми та черевцем. Найскладнішу будову мають тимпанальні органи коників (рис. 144). Барабанна перетинка в них ізсередини
щільно притиснута до двох широких трахей, зверху головної з них розміщені сколопідії, зібрані в три групи: підколінний орган, проміжний орган і слуховий гребінь. Коливання барабанної перетинки спочатку передаються на трахейний стовбур, а від нього — до сколопідіїв.
Рис. 144. Тимпанальний орган коника:
а - загальний вигляд; б - поздовжній зріз; 1 - отвір тимпанального органа; 2 - головна трахея; 5-слуховий нерв; 6 - проміжний орган; 5 - слуховий гребінь; 6 - підколінний орган; 7- нерв
Терморецепторами комах є трихоїдні, базиконічні та целоконічні сенсили, розміщені на різних частинах тіла, в основному на антенах. Одні з них сприймають лише холод (зниження температури), це трихощні сенсили, інші —тепло (підвищення температури) — базиконічні та целоконічні.
Гігрорецептори представлені базиконічними або целоконічними сенсилами, розташованими на антенах. Одна й та сама сенсила може бути і терморецептором, і гігрорецептором, якщо вона має кілька чутливих клітин.
Хеморецепторні сенсили комах можна поділити на дві групи: нюхові, або дистантні, які сприймають молекули летючих речовин у дуже малих концентраціях, та смакові, або контактні, що сприймають хімічний стимул у контакті з речовиною. Вони трапляються на різних придатках тіла: антенах, ротових кінцівках, лапках ніг, церках, яйцекладі.
Кутикулярні частини хеморецепторних сенсил різноманітні за формою, але мають характерні риси ультраструктури —наявність однієї або кількох пор на кінчику чи на всій поверхні кутикулярної частини сенсили (рис. 145). Через ці пори молекули хімічних речовин можуть вільно проникати до рецепторної поверхні нейронів. Рецепторами смаку найчастіше бувають трихощні сенсили, нюху — трихощні, базиконічні, целоконічні, плакощні та деякі інші. Друга особливість — це наявність у сенсили не однієї, а кількох чутливих клітин, кожна з яких реагує на подразнення певного групою речовин. Так, до складу рецепторів смаку, як правило, входять 3 або 4 чутливі клітини. Одна з них реагує на розчини солей, друга збуджується розчинами цукрів, третя — чистою водою. До складу деяких дистантних хеморецепторів входить більше десяти чутливих клітин.
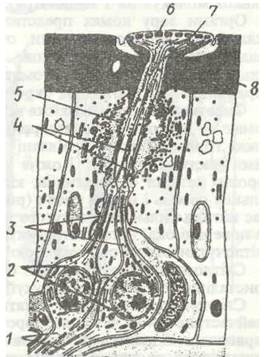
Рис. 145. Схема будови хеморецепторної сенсили комах:
1 - аксон чутливої клітини; 2 - чутлива клітина; 3 - дендрит; 4 - видозмінений джгутик; 5 - кутикулирна трубочка, що фіксує джгутик; 6 - кутикулярна частина; 7 - пори; 8 - кутикула
Комахи мають надзвичайно розвинений нюх, особливо на специфічні речовини, що їх приваблюють (атрактанти) або викликають відразу (репеленти). Серед атрактантів особливо важливе значення в житті комах мають харчові й статеві. Перші полегшують їм пошук їжі, другі, які виділяються самицями, допомагають самцям знаходити їх на великій відстані. Відомо, наприклад, що незапліднені самиці деяких метеликів здатні приваблювати самців на відстані 3—9 км; це пояснюється надзвичайною чутливістю нюхових сенсил самців: вони здатні реагувати на статевий атрактант на великій відстані та при дуже малій його концентрації (до кількох молекул на 1 м3 повітря).
Органи зору комах представлені трьома типами очей: складними, або фасетковими, очима, латеральними та дорзальними вічками. Фасеткове око складається з великої кількості фоторецепторів — оматидіїв, а кожне латеральне та дорзальне вічко відповідає окремому фоторецептору.
Фасеткові очі мають майже всі дорослі комахи та личинки комах із неповним перетворенням. Вони розташовані по боках голови й тісно пов'язані з добре розвиненими зоровими частками мозку. Кожне око складається з окремих зорових сенсил — оматидіїв, кількість яких може досягати кількох сотень і навіть тисяч (рис. 146). Оматидій (рис. 147) має вигляд сильно витягнутого конуса, зверненого основою до поверхні ока, і становить сукупність світлозаломлюючого, світлочутливого та світлоізолюючого елементів.
Світлозаломлюючий апарат складається з кришталика та кристалічного конуса, які в сукупності виконують роль лінзи.
Світлочутливий апарат оматидія складається з кількох (найчастіше 8—9) чутливих зорових (ретинальних) клітин із нервовими відростками, які зв'язують їх із мозком. Вони розташовані по колу, подібно до часток апельсина. Кожна зорова клітина має по всій довжині внутрішньої поверхні особливу структуру складної мікроскопічної будови -рабдомер. Кожен рабдомер складається з великої кількості мікроворсинок, де знаходяться зорові пігменти (рис. 147, в). Рабдомери всіх зорових клітин щільно прилягають один до одного й утворюють зорову паличку, аборабдом. Саме в раб- домах здійснюється фоторецепція — перетворення світлового сигналу на нервовий імпульс.
Світлоізолюючий апарат оматидія складається з кількох пігментних клітин, які оточують кристалічний конус і ретинулу оматидія. Вони ізолюють ці утвори у суміжних оматидіїв, запобігаючи проникненню променів світла до рабдомів через сусідні оматидії. У денних комах пігмент розміщений рівномірно по всій довжині пігментних клітин і цілком ізолює оматидій від сусідніх (опозиційне око}. У нічних комах, які активні при дуже слабкому освітленні, пігмент здатний переміщуватись і нагромаджуватись лише у верхніх частинах пігментних клітин (супер- позиційне око). Завдяки цьому промені світла потрапляють на рабдоми не одного, а кількох сусідніх оматидіїв, що істотно підвищує чутливість ока до світла. Крім того, в очах цього типу рабдом укорочений і лежить у нижній частиш оматидія.
Денні комахи, як уже зазначалося, мають апозиційний зір. Завдяки оптичній ізоляції за допомогою пігментних клітин кожен оматидій перетворений на ізольовану тонку трубку, тому до нього можуть доходити тільки ті промені, які йдуть через кришталик та збігаються з поздовжньою віссю оматидія. Вони й досягають рабдома. Отже, поле зору кожного оматидія дуже мале й сприймає лише дуже малу частину предмета, який розглядається. Проте велика кількість оматидіїв дає змогу різко збільшити поле зору взаємним прикладанням (апозицією) одного до одного. В результаті з окремих найменших частин складається, як у мозаїці, єдине загальне зображення.
Рис. 146. Схема будови фасеткового ока комахи:
1 - рогівка (прозора кутикула); 2 - кришталевий конус; 3 - пігмент між оматидиши
Комахи мають кольоровий зір. Найдосконаліший він у бджолиних і денних метеликів. Проте в комах, на відміну від людини, видима частина спектра захоплює також зону ультрафіолету (короткі хвилі); навпаки, довгохвильова частина його коротша й закінчується на оранжово-червоному, не доходячи до червоного.
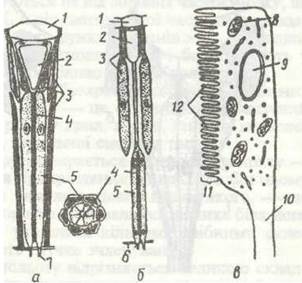
Рис. 147. Схема будови оматидія:
а - денних комах; б - нічних; в - окрема світлочутлива клітина;
1 - кришталик; 2 - кришталевий конус; 3 - пігментні клітини; 4 - рабдом; 5 - чутливі клітини; б ~ базальна мембрана; 7 - аксони чутливих клітин; 8 - мітохондрія; 9 - ядро; 10 - аксон; 11 — рабдомер; 12 — мікроворсинки
Комахи мають унікальну здатність до сприйняття поляризації світла. Денне світло поляризоване, проте людина не здатна сприймати поляризацію. Комахи, завдяки такій здатності, дістають змогу орієнтуватися по небу навіть тоді, коли воно затягнуте хмарами (астронавігація). Механізм цього явища полягає в упорядкованому розташуванні мікроворсинок у рабдомі.
«
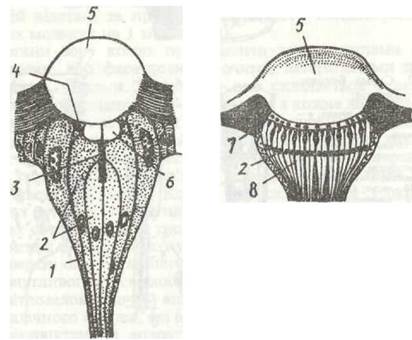
Рис. 148. Схема будови латерального (я) та дорзального (б) вічок комах:
1 - мантійна клітина; 2 - зорові клітини; 3 - рабдом; 4 - корнеагенні клітини; 5 - рогівка (кришталик); 6 - кристалічне діло; 7 - пігментована гіподерма; 8 - пігментні клітини сітківки
Латеральні (бічні) вічка трапляються в личинок комах із повним перетворенням, а також в окремих представників імаго, наприклад, бліх, самців червеців. Ці вічка містяться по боках голови в кількості від одного до 30 з кожного боку. При переході комахи в дорослий стан вони руйнуються й замінюються на фасеткові очі.
Будова латеральних вічок (рис. 148, а) різноманітна: в одних комах, наприклад у гусені метеликів, вони нагадують окремий оматидій, у інших — близькі за будовою до дорзальних вічок (личинки пильщиків).
Дорзальні вічка (рис. 148, б) бувають у дорослих комах (як правило, в таких, які добре літають) та в личинок комах із неповним перетворенням. Найчастіше три дорзальних вічка розташовані у вигляді трикутника на верхній поверхні голови. Вічко має одну лінзу —двоопукле потовщення прозорої кутикули; під нею лежить тонкий шар клітин, який і виділяє лінзу, а під ним залягає сітківка (рис. 148, б). Вона складається з окремих світлочутливих і пігментних клітин. По периферії сітківки розташовані гіподермальні пігментні клітини, які захищають вічко від бокових променів світла. Дорзальні вічка іннервуються не від зорових часток мозку, як фасеткові та латеральні, а від центральної частини протоцеребрума. Вони не виконують функції органів зору, але підвищують світлочутливість фасеткових очей і беруть участь у зоровій орієнтації комах, особливо під час польоту.
Комахи — єдина група безхребетних тварин, здатних активно літати. Політ комах — це складний фізіологічний процес, який включає роботу крил, м'язів, гангліїв нервової системи, органів чуття, дихальної системи тощо.
Крило комахи при русі опирається на виріст плейриту — стовпчик поблизу місця прикріплення крила. Завдяки цьому крило стає двоплечим важелем, довге плече якого — це крилова пластинка, а коротке —невеличка ділянка біля його основи. Основа крила укріплена кількома дрібними склеритами, які забезпечують гнучке зчленування.
Рух крила під час польоту відрізняється великою складністю. Кожне крило може розглядатись як своєрідний пропелер, що створює поступальний рух комахи. У нерухомо закріпленої комахи крило описує складну фігуру, схожу на вісімку, причому його нахил (кут атаки) весь час змінюється. Крім того, під час руху крила створюється також підйомний ефект, коли воно опускається вниз у вигляді горизонтальної площини. Висока частота помахів забезпечує поєднання підйомного й поступального ефектів, що й дає змогу комасі летіти вперед, стійко утримуючись у повітрі.
Рух крил під час польоту забезпечується координованою роботою системи м'язів непрямої та прямої дії. Для більшості комах головну роль відіграють м'язи непрямої дії. Вони безпосередньо до крила не прикріплюються, а сполучені з різними склеритами грудей (рис. 149). Це дві групи антагоністичних м'язів. Перша з них, дорзовентральні м'язи, прикріплюєгься одним кінцем до тергіта, другим — до стерніта й основи ніг. У момент скорочення вони опускають тергіт, який тисне на основу крила, підіймаючи його вгору. Друга група, поздовжні м'язи, розташована уздовж спинки грудей; вони кріпляться до фрагм тергітів і під час скорочення зближують фрагми; при цьому спинка вигинається догори, а крила опускаються. Так забезпечується поступальний рух. Важливе значення тут має пружність кутикули тергітів, які після скорочення м'язів набувають початкової форми.
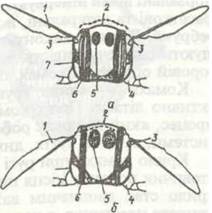
Рис. 149. Схема роботи крил ко мамі:
а — стадія піднятого крила; б — стадія опущеного крила; 1 - крило; 2 - тергет; 3 - стовпчик; 4 - основа нош; 5 - поздовжній м'яз;
6 - дорзоаентральннй м'яз; 7 - плейрит
М'язи прямої дії приєднані безпосередньо до основи крила та склеритів грудей. З їх допомогою крило повертається вздовж поздовжньої осі, змінюючи кут нахилу, а також відводиться вперед чи назад, регулюючи швидкість та напрямок польоту. Лише в бабок (ряд Odonata) у польоті майже виключно беруть участь м'язи прямої дії.
Зрозуміло, що ліве й праве крила обох пар діють синхронно, причому в більшості комах у польоті беруть участь або задні (коли передні крила перетворені в надарила), або передні (у двокрилих) крила.
Часто під час польоту передні та задні крила зчіплюються й діють як єдине ціле (метелики, перетинчастокрилі). У бабок кожне крило діє автономно, чим досягається надзвичайна швидкість і маневровість польоту.
Однією з найдосконаліших форм польоту є так званий стоячий політ. Він характерний для деяких перетинчастокрилих, метеликів та мух і полягає в тому, що комаха «нерухомо» тримається в повітрі в одній точці. Наприклад, метелик-бражник, зависаючи над квіткою, запускає в неї свій хоботок і живиться нектаром.
Швидкість польоту в різних комах різна: у кімнатної мухи вона не перевищує 8 км/год, у сарани — 12,6, у джмеля — 18, у бражника — 54, а у деяких бабок — навіть до 100 км/год.
Великі комахи, поверхня крил яких значна, здатні досягати великої швидкості польоту при невеликій частоті помахів крил. Дрібні ж форми з маленькими крилами збільшують швидкість польоту, підвищуючи частоту биття крил. Кількість помахів крила за секунду коливається від 5—10 (денні метелики) до кількох сотень у двокрилих, а у комарів-дзвінців (родина Chironomidae) та дрібних комарів-мокреців (родина Ceratopogonidae) — до 1000 помахів.
У комах із досконалим польотом (бабки, перетинчастокрилі, деякі двокрилі та інші) на передньому краї крила жилки утворюють потовщення — птеростигму. Вона при великих швидкостях гасить шкідливу крилову вібрацію, яка в аеродинаміці називається флатером. У добрих літунів поздовжні жилки частково зливаються та переміщуються до переднього краю крила. Цікаво, що в сучасних літаках для запобігання вібрації на перед ньому краї крила встановлюють додатковий вантаж.
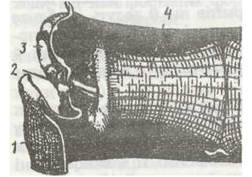
Рис. 150. Звуковий (тимбальний) орган цикади
Роботою крилевих м'язів керують ганглії відповідних грудних сегментів. У грудних гангліях знайдено окремі нейрони, здатні у відповідь на сигнал з головного мозку генерувати ритмічні нервові імпульси з певного постійною частотою, спричиняючи скорочення м'язів. Отже, мозок не контролює кожен рух крила; грудні ганглії здатні самостійно підтримувати прямолінійний політ.
Частота помахів крил відповідає частоті скорочень крилових м'язів. Як уже зазначалося, м'язи, що забезпечують політ, можуть бути синхронними й асинхронними. Синхронні м'язи забезпечують політ комах з порівняно низькою частотою биття крил (сарана, метелики, деякі жуки). У них один нервовий імпульс із грудного ганглія спричиняє одне скорочення м'язів і відповідно один цикл (підняття та опускання) роботи крил. Асинхронні, або швидкі, м'язи властиві комахам із високою частотою биття крил (перетинчастокрилі, двокрилі). На один нервовий імпульс вони відповідають кількома (від 2—3 до 10—15) скороченнями, при цьому один нервовий імпульс викликає автоколивання м’яза; коли вони затухають, новий імпульс підтримує їх. Цьому сприяє особлива пружність кутикули.
Здатність крилевих м'язів скорочуватися з частотами, незвичайними для м'язів інших тварин, забезпечується структурними особливостями самих м'язів (вони містять дуже великі міофібрили й тому звуться фібрилярними) та інтенсивним постачанням їх киснем. Найдрібніші розгалуження трахей — трахеоли — входять усередину м'язових волоконець і утворюють там складну систему внутрішньоклітинних розгалужень. Здатність до швидких скорочень забезпечується високим рівнем метаболізму крилових м'язів, особливостями температурного режиму їхньої роботи (при цьому температура підвищується до 40—42°С), а також вдалим використанням еластичних властивостей кутикули грудних сегментів та самих м'язів.
Велике значення в стимуляції початку польоту та його підтриманні мають органи чуття. Механорецептори ніг сигналізують про відрив тіла комахи від субстрату, що є необхідною умовою початку польоту; вітрочутливі сенсили на голові сигналізують нервовій системі про швидкість й напрямок польоту (подібні рецептори є ще в основі крил та на їхній поверхні) тощо. Велика роль у підтриманні польоту й маневруванні належить джонстоновому органу, що міститься в антенах, а також очам, які визначають швидкість польоту й положення тіла в просторі. У двокрилих важливі функції при запуску польоту й стабілізації тіла в просторі виконують дзижчальця.
Комахи характеризуються складними формами поведінки. Перш за все це стосується комплексу інстинктів турботи про нащадків. У межах класу Insecta можна знайти всі переходи від найпростіших форм до найскладніших, властивих суспільним комахам.
У найпростіших випадках турбота про нащадків виявляється у відкладанні самицями яєць біля харчового субстрату личинок або просто в нього (більшість метеликів, мухи, жуки та ін.). Складнішою формою є створення запасів їжі для потомства та побудова тих чи інших спеціальних укриттів, де розвивається молодь. Найбільш поширене це явище у перетинчастокрилих.
Наприклад, одиночні бджоли будують гнізда (в грунті, деревині або іншому субстраті) з низки комірок, причому в кожну з них умішують запас нектару й пилку рослин, після чого ці комірки запечатують. Риючі оси запасають у нірках паралізованих ними комах та відкладають на них яйце. Личинка, що вийшла з яйця, поїдає живу, але нерухому жертву. Для того щоб паралізувати здобич, оса проколює своїм жалом її тіло й уражує ганглії черевного нервового ланцюжка. Більшість видів при цьому спеціалізується на пошуках певного виду здобичі: як правило, оси амофіли паралізують гусінь совок та п'ядунів, сфекси — саранових та коників, церцеріси — деяких жуків тощо. Пошук, паралізація або умертвіння здобичі та всі наступні дії цих ос супроводжуються складною поведінкою.
Найскладніша поведінка у суспільних комах, які живуть великими сім'ями. Це терміти, деякі бджоли, оси, джмелі та мурашки. Для цих комах характерний поліморфізм, тобто диференціювання особин сім'ї на кілька форм (каст): самців, самиць (матка, «цариця»), робітників. Часто бувають ще різні форми робітників, спеціалізовані на виконанні певних функцій, наприклад солдати, які захищають гніздо від ворогів, фуражири та ін.
Робочі особини годують личинок, добувають їжу, чистять та захищають житло, а статеві призначені лише для розмноження.
Поліморфізм виникає завдяки виділенню маткою особливих біологічно активних речовин — телергонів, які впливають на поведінку робочих особин і гальмують розвиток у них статевих органів. Отже, робітники й солдати — це неплідні самиці і самці. Важливе значення має й спрямоване виховання личинок —посилене спеціальне годування тих, з яких вийдуть самиці, та обмежене годування майбутніх робітників.
У суспільних комах форми поведінки, пов'язаної з турботою про нащадків, найбільш різноманітні. Це виявляється в постійному догляді за личинками, їх годівлі, охороні, а інколи й викраданні личинок з інших родин (у деяких мурашок). Деякі види мурашок і термітів ще й ведуть особливе «господарство»: мурашки охороняють і навіть розводять попелиць, від яких одержують солодкі виділення; терміти й мурашки-листогризи з роду Atta вирощують у своїх гніздах гриби, гіфами яких живляться.
Усі зазначені форми поведінки не потребують навчання: це генетично закріплені інстинкти. Проте велику роль у поведінці комах, особливо суспільних, відіграють також умовні рефлекси. Вирушаючи за здобиччю, ці комахи здатні запам'ятовувати місцезнаходження джерела їжі, дорогу до нього й назад, до гнізда. Медоносних бджіл можна навчити прилітати на той чи інший кольоровий субстрат або намальовану на ньому фігуру. Навіть у таких одиночних комах, як таргани, можна виробити умовні рефлекси.
Велике значення для комах, навіть одиночних, мають засоби спілкування, за допомогою яких відбувається обмін певного інформацією. Вони дуже різноманітні у різних комах. Це звукові сигнали, пахучі речовини, особливі біологічно активні речовини, злизувані комахами, світлові сполохи, особлива форма поведінки, яку сприймають інші особини.
Частина комах здатна створювати спеціальними органами звуки певних частот. У більшості це дві поверхні, одна з яких пильчаста, а інша має виступ або ребро. Від тертя цих поверхонь виникає звук. Наприклад, у коників та цвіркунів ліве переднє крило частково лежить на правому. На одному з них поперечна жилка при основі крила потовщена і зубчаста («напилок»), на другому, проти «напилка», гостре ребро. При цвіркотанні тварина швидко вібрує крилами. Саранові на задньому стегні мають ряд зубців («смичок»), якими труть по спеціалізованій потовщеній жилці переднього крила.
У цикад органи звуку розташовані в заглибленнях біля основи червця; це мембрана, яка швидко коливається під дією спеціальних м'язів (рис. 150). Звуки цих комах часто сильні й мелодійні: деяких цикад у Давньому Римі тримали в клітках замість співочих птахів. Дзижчальця багатьох двокрилих під час роботи спричинюють звуки певної частоти, які сприймаються іншими особинами того самого виду. «Співають» переважно самці, які таким чином приваблюють самиць, інколи (двокрилі, вовчки) звуки можуть утворювати обидві статі. Звуки комах певного виду можуть приваблювати їхніх ворогів. У Північній Америці самиці одного виду мух родини Sarcophagidae приваблюються співом самців цикад, де й відкладають паразитичні личинки. Дослідники записали спів цикади на магнітофсії і ввімкнули його в природних умовах: за дві години біля нього зібралось кілька сотень самиць мухи, готових до відкладення личинок.
Відповідно до розвитку звукових органів у цих комах розвинені й органи слуху. Певні види комах пристосовані до сприйняття лише тих звуків, які мали біологічного значення й не реагують на «сторонні» звуки. За допомогою звукових сигналів забезпечується зустріч особин протилежної статі, підтримується репродуктивна ізоляція у близьких видів, регулюються внутріпопуляційні та родинні відносини.
Велике значення у спілкуванні комах мають особливі біологічно активні речовини — телергони, за допомогою яких здійснюється спілкування з особинами свого (феромони) або інших (гетеротелергони) видів.
Серед феромонів розрізняють кілька груп. Статеві феромони виділяються самицями для приваблення самців. Найкраще їх вивчено у лускокрилих; залози, що їх виділяють, розташовані поблизу кінця черевця. Запах феромону самець може відчувати на відстані в кілька кілометрів.
Інколи феромони приваблюють особин обох статей, і внаслідок утворюються їх скупчення (феромони агрегації). Наприклад, феромон клопа шкідливої черепашки, який виділяється грудними залозами, спричиняє скупчення цих тварин на кормових рослинах — злакових.
У робочих мурашок і термітів на кінці черевця є залоза, виділеннями якої мітиться шлях (слідові феромони). Це полегшує повернення в гніздо, а також показує дорогу іншим особинам до поживи. Так, більшість мурашок залишає пахучі сили вздовж «мурашиних доріг». Слідові феромони мають вузьку видову специфічність: мурашки прямують по пахучих слідах лише свого виду, не повертаючи на сліди мурашок інших видів, які багаторазово перехрещуються з їхніми слідами. Деякі види мурашок за допомогою слідових феромонів не просто стимулюють робочі особини рухатись уздовж них, а й передають інформацію про напрямок і кількість їжі, яка є в тому місці, куди веде слід. У різних комах слідові феромони секретуються різними залозами: у термітів вони відкриваються на четвертому етерніті черевця; у мурашок у різних частинах тіла містяться залози, що виробляють різноманітні феромони; бджоли мають спеціальну насонову залозу, яка міститься на спинній стороні між V та VI сегментами черевця; пахучий секрет цієї залози приваблює інших бджіл. Бджоли залишають пахучу мітку на квітах, з яких збирають нектар, і це є додатковим орієнтиром для інших особин.
У загрозливій ситуації комаха виділяє феромон тривоги. Реакція інших особин виду може бути різною: захист, збудження, мобілізація, уникнення небезпеки тощо. Ці реакції характерні переважно для суспільних комах.
У суспільних комах —термітів, мурашок, бджіл —велике значення мають феромони стабільності структури сім’ї. Мандибулярні залози бджолиної матки виділяють маслянисту речовину зі специфічним запахом, яка змащує тоненькою плівкою все її тіло. Це так звана маточкова речовина, яку робочі особини злизують або сприймають на запах; вона свідчить про наявність матки в сім'ї. Ця речовина гальмує розвиток яєчників у робочих особин. Постійне злизування цього телергону з тіла матки та розповсюдження його серед усіх бджіл у вулику — обов'язкова умова нормального існування бджолиної сім'ї. Коли матка гине й, отже, припиняється виділення маточкової речовини, робітники вигодовують з личинок нових маток; у деяких із них самих навіть починають розвиватись яєчники. Запах маточкової речовини приваблює й самців під час парування. У мурашок і термітів мандибулярними залозами теж виділяється спеціальна речовина, яку злизують робочі особини. Так у цих комах відбувається саморегуляція співвідношення між кастами в колонії.
Деякі комахи виділяють речовини, що впливають на поведінку особин не свого, а інших видів (гетеротелергони). У мірмекофільних жуків (тобто тих, які мешкають у мурашниках) родів Atemeles та Lomechusa (родина Staphylinidae) на черевці є залози, які продукують речовини, що надзвичайно приваблюють мурашок. Ці речовини звуться ліхневмонами. Коли такий жук потрапляє до мурашника, то мурашки весь час облизують його, дбають про нащадків жука й навіть згодовують їм свої яйця. Поведінка мурашок під наркотичним впливом ліхневмонів змінюється. Вони занедбують мурашник, не годують нормально личинок, з яких в результаті замість самиць і самців виходять недорозвинені особини, нездатні до розмноження, й згодом мурашник гине.
Найскладнішою формою передавання інформації у комах можна вважати високоспеціалізовані рухові реакції — «ганці». Відомо, що бджола-розвідниця, повернувшись у вулик, починає танцювати на сотах, описуючи при цьому певні фігури. За допомогою танців вона передає іншим бджолам інформацію про напрямок, відстань до місця взятку й навіть про кількість корму. Бджоли, сприймаючи ці сигнали, знаходять у природі джерело корму.
Багато видів комах здатні в певних ситуаціях застосовувати речовини, спрямовані проти інших істот. Найчастіше це засоби хімічного захисту від ворогів, іноді — нападу на здобич.
Наприклад, самиці жалячих перетинчастокрилих (оси, бджоли, джмелі, деякі мурашки) мають знаряддя захисту — жало. Це видозмінений яйцеклад, зв'язаний з отруйною залозою, їхня отрута небезпечна й для людини: 3 — 4 уколи великої оси — шершня (Vespa crabro) можуть спричинити смерть, особливо людей, які страждають алергією на отруту перетинчастокрилих. У деяких мурашок (Formica та інших) жало редуковане, і вони вибризкують отруту (розчин мурашиного альдегіду) на нападника або прокушують його покриви жувальцями, а потім упорскують отруту в ранку.
Риючі оси, про яких уже згадувалося, паралізують інших членистоногих уколом жала в ганглії нервової системи, а потім відкладають на жертві яйця.
Отруйні залози, що відлякують ворогів, має гусінь багатьох метеликів. Вони розташовані біля основи зазубрених щетинок, які легко відламуються. В Україні особливо небезпечна отрута гусені золотогузки (Euproctis chrysorrhoea), яка у людини призводить до сильних шкірних алергічних реакцій. Клопи-щитники виділяють секрет грудних залоз, який спричиняє параліч і смерть хижих комах, відлякує птахів та інших комахоїдних тварин неприємним запахом.
У багатьох комах отруйна гемолімфа. Серед них найбільш відомі жуки-наривники (родина Meloidae), гемолімфа яких містить отруйну речовину кантаридін. Якщо такого жука потурбувати, він виділяє краплини гемолімфи через отвори, розташовані між стегнами та гомілками ніг. Отрута відлякує павуків, хижих комах, птахів. Навіть на великих тварин вона
діє згубно. Верблюд чи кінь, випадково проковтнувши з травою наривника, захворюють на запалення кишечника і часто гинуть. У людини отрута наривників викликає, опіки. Виділяють отруйну гемолімфу й інші комахи: жуки-сонечка, личинки тополевого та осикового листоїдів та деякі інші.
Одним із найскладніших є пристосування для відлякування ворогів у жуків-бомбардирів (рід Brachinus, родина Carabidae). Вони стріляють рідиною, яка, немов снаряд із
мініатюрної гармати, вилітає ззаду із черевця жука й пере- творюється на димовий клубочок. У цих жуків біля анального отвору є дві залози, що сполучаються між собою. В одній залозі міститься 10 %-й гідрохінон та 25 %-й перекис водню, а в другій — фермент каталаза, яка розкладає перекис водню на кисень і воду, та фермент пероксидаза, що перетворює гідрохінон на хінон із виділенням тепла. При викиданні назовні ця гаряча (до 100 °С) маса вибухає з гучним тріском та утворенням їдкого диму, відлякуючи ворогів.
Комахи розмножуються лише статевим шляхом. Як правило, вони роздільностатеві й часто мають чіткий статевий диморфізм, який виявляється в розмірах тіла, забарвленні, розмірах вусиків тощо. Інколи особливо різко, наприклад у самця жука-оленя, розвиваються величезні мандибули, а в жука-носорога — ріг. Проте часто обидві статі майже однакові, й розрізнити їх можна лише за будовою статевих придатків (геніталій). У деяких комах (наприклад, попелиці) має місце партеногенез.
Статева система самиці складається з пари яєчників, пари яйцепроводів, непарного яйцепроводу, придаткових статевих залоз, сім'яприймача та часто яйцекладу (рис. 151). Кожен яєчник складається з яйцевих трубок, яких у різних комах може бути від 1—4 до 100, а інколи й більше, наприклад, у термітів їх більше ніж 2400.
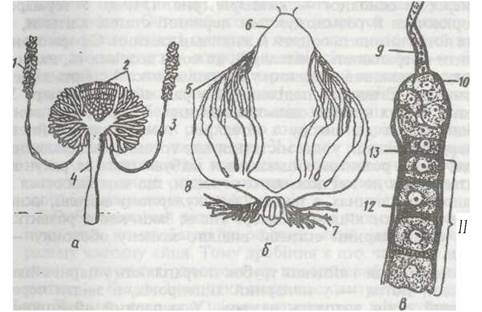
Рис. 151. Статева система таргана
Кожна яйцева трубка складається з верхівкової частини — гермарія та основної — вітелярія (рис. 151, в). У гермарії утворюються й розмножуються первинні статеві клітини, з яких потім виходять ооцити й живильні клітини. Сформовані ооцити потрапляють у вітелярій, де вони дозрівають, нагромаджують поживні речовини у вигляді жовтка й стають яйцеклітинами. Вітелярій поділений на ряд яйцевих камер. У кожній із них є лише одна яйцеклітина. В міру росту вона наближається до виходу з вітелярія, тому останні яйцеві камери найбільші; у них містяться вже готові до відкладання яйця. Ріст і розвиток яйцеклітини відбуваються за рахунок надходження до неї поживних речовин, що виробляються в живильних клітинах, а також у фолікулярному епітелії, який формує стінки яйцевих камер. Після закінчення розвитку яйця фолікулярний епітелій виділяє захисну оболонку хиріші.
Дозрілі яйця з яйцевих трубок потрапляють у парні яйцепроводи, потім — у непарний яйцепровід, а звідти через статевий отвір виходять назовні. У непарний яйцепровід впадає протока сім'яприймача, призначеного для зберігання сперматозоїдів, які потрапляють до нього під час парування. Яйце запліднюється, проходячи через непарний яйцепровід під час відкладання яєць; сперматозоїди в цей час виходять із сім'яприймача й проникають у яйце. У багатьох комах у непарний яйцепровід відкривається мускуляста парувальна сумка; при копуляції сперма спочатку потрапляє в цю сумку, а потім — у сім'яприймач. До непарного яйцепроводу відкриваються також придаткові статеві залози, які виконують різні функції — виділяють секрет для приклеювання яєць до субстрату, для утворення навколо групи яєць захисної оболонки — оотеки (таргани, богомоли) або ворочка (саранові). Часто із статевим отвором зв'язаний яйцеклад для відкладання яєць у більш-менш твердий субстрат (ґрунт, тканини рослин чи тварин тощо).
Статева система самця складається з пари сім'яників, пари сім'япроводів, сім'явипорскувального каналу, придаткових статевих залоз і копулятивного органа. Сперматозоїди з сім'яників потрапляють до сім'япроводів, з них — у сім'явипорскувальний канал, і при паруванні виводяться назовні. Часто копулятивний орган утворює зовнішні статеві придатки — геніталії, будова яких має видову специфічність і використовується в систематиці комах. Придаткові залози, які відкриваються у сім'явипорскувальний канал, у багатьох комах призначені для вироблення сперматофору, який уводиться в статевий отвір самиці чи прикріплюється до нього.
Розмноження комах регулюється гормональною системою. Нейросекреторні клітини мозку самиці активізують діяльність прилеглих тіл, які виділяють у гемолімфу ювенільний гормон. Останній стимулює розвиток яєчників. Механізм цього явища полягає в тому, що ювенільний гормон стимулює синтез у жировому тілі вітелогенінів — специфічних білків, які надходять через стінки яєчників до ооцитів і перетворюються там на жовток. Активність нейросекреторних клітин мозку залежить від зовнішніх факторів (температури, довжини світлового дня) та внутрішніх (стан яєчників, живлення, парування). У самців процеси сперматогенезу не підлягають дії гормонів; у них гормональні фактори регулюють лише статеву активність, розвиток придаткових залоз і формування сперматофорів.
Ембріональний розвиток комах має свої особливості. Яйцеклітини комах багаті на жовток, який займає всю центральну частину яйця. Тому дробіння в них часткове, поверхневе; у результаті утворюється поверхневий шар однакових клітин — бластодерма, яка покриває центральну масу жовтка. Частина ядер при цьому залишається в жовтковій масі; пізніше за їх рахунок утворюються жовткові клітини. На майбутній черевній стороні зародка клітини бластодерми починають інтенсивніше ділитися, бластодерма потовщується. Ця ділянка зветься зародковою смужкою. Пізніше за її рахунок формується більша частина зародка.
У зародковій смужці відбувається гаструляція (найчастіше інвагінацією або епіболією), причому спочатку утворюються лише два зародкових листки: ектодерма та мезодерма. Ектодерма дає початок зовнішнім покривам тіла, трахеям, нервовій системі, статевим протокам, передньому та задньому від ділам кишечника, що виникають як впинання покривів на передньому та задньому кінцях зародка.
Із мезодерми утворюється мускулатура, жирове тіло, серце з аортою, гемоцити. Мезодерма бере участь також у розвитку статевої системи. Статевий зачаток, як правило, відокремлюється значно раніше від інших тканин у вигляді парної групи клітин біля заднього кінця зародка.
Ентодерма, з якої походить середня кишка, у різних комах може будуватися двома шляхами: або за рахунок ядер, що залишилися в товщі жовтка й мігрували до зародкової смужки, або з невеличких зачатків на дні передньої та задньої кишок, які розростаються назустріч один одному, доки не утвориться суцільна трубка — кишечник.
Водночас із диференціацією зародкових листків над зародком виникають дві оболонки — зовнішня, або сероза, та внутрішня, або амніон. Зародок опиняється під захистом цих оболонок, що забезпечують надійніші умови для нормального розвитку ембріона (рис. 152).
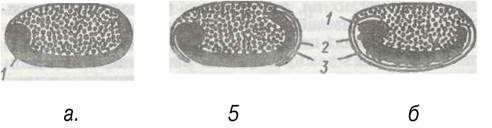
Рис. 152. Один Із способів утворення зародкових оболонок у комах:
а - зародок перед утворенням оболонок; б - початок й утворення; в - завершення; 1 - головний кінець зародка; 2 - сероза; 3 — амніон
Паралельно з розростанням зародка в оболонках починається його сегментація. На головному кінці з'являються зачатки очей, відокремлюються сегменти голови, грудей та черевця. З'являються зачатки кінцівок, у тому числі й на черевці. Пізніше черевні кінцівки зникають. Мезодерма при цьому теж сегментується на кілька целомічних мішечків, які потім розпадаються.
Урешті-решт увесь жовток вичерпується, і сформований зародок заповнює все яйце. Він прогризає чи прориває оболонку яйця й виходить назовні.
Постембріональний розвиток комах відбувається з метаморфозом і має епіморфний характер. Це означає, що з яйця виходить личинка з усіма сегментами, яка більш-менш відрізняється від дорослої особини — імаго. Розрізняють два основні типи метаморфозу — неповне та повне перетворення.
При неповному перетворенні, або гемі/метаболії (рис. 153), комаха проходить такі фази розвитку: яйце, личинка, імаго. З яйця виходить личинка, зовні схожа на дорослу особину. Вона має фасеткові очі, такий самий які в дорослих ротовий апарат, а на старших стадіях — зовнішні зачатки крил. Спосіб життя личинок багатьох видів з неповним перетворенням і дорослих комах однаковий. Ці личинки звуться німфами. Окрему модифікацію становлять так звані наяди — личинки одноденок, бабок і веснянок. Вони теж схожі на імаго, мають фасеткові очі й на старших стадіях — зачатки крил, але живуть у воді й мають спеціальні личинкові органи — зябра, маску (личинки бабок) тощо.
Рис. 153. Неповний метаморфоз прусака Blattella germanica:
а - яйце; б-е - шість личинкових стадій (німфи); ж — імаго
Ріст і розвиток личинки супроводжуються періодичними линяннями, кількість яких, як правило, стала для певних таксонів (найчастіше родин). З кожним линянням личинка за розмірами, пропорціями тіла, розвитком крил наближається до дорослої фази; під час останнього линяння вона перетворюється на імаго.
При повному перетворенні, або голометаболії, комахи проходять такі фази: яйце, личинка, лялечка, імаго. З яйця виходить личинка, яка різко відрізняється від імаго більшою гомономністю метамерії, відсутністю зовнішніх зачатків крил, слабкішим розвитком органів чуття. Вона не має фасеткових очей, антени її значно коротші; часто ротовий апарат у неї інший, ніж у дорослої комахи (наприклад, у гусені метеликів — гризучий, а в імаго — сисний). Личинки комах із повним перетворенням живуть в інших умовах, ніж дорослі, і з ними не зустрічаються. Більшість органів личинки має тимчасовий характер, наприклад черевні ноги в личинок метеликів та пильщиків, шовковидільні залози у гусені метеликів, личинок деяких ос, їздців, пильщиків тощо.
Личинки комах із повним перетворенням дуже різноманітні (рис. 154). Серед них трапляються червоподібні, малорухомі, з м'якими покривами, інколи навіть без голови (личинки мух), часто безногі або зі слабко розвиненими грудними ногами (личинки жуків, перетинчастокрилих). Інші (гусенеподібні) більш рухомі, з трьома парами грудних і 2—8 парами коротеньких черевних ніг (гусінь метеликів, личинки пильщиків). Рідше трапляються рухомі личинки з твердими покривами, добре розвиненими грудними кінцівками та ротовим апаратом із гострими щелепами. Це хижі личинки жуків-турунів, плавунців, сітчастокрилих.
Рис. 154. Личинки комах із повним метаморфозом:
а - травневого хруща; б - мухи; в - метелика (гусінь); г - бджоли; д - жука туруна (камподеоподібна личинка)
Личинкові ознаки зберігаються під час усієї личинкової фази. Із кожним линянням личинка лише збільшується в розмірах, а необхідні для метаморфозу зміни відсуваються до останнього личинкового линяння. Закінчивши свій ріст, личинка останньої стадії припиняє живлення, стає нерухомою, линяє востаннє й перетворюється на лялечку.
Характерна особливість лялечки — нерухомість (за деякими винятками) і нездатність до живлення (вона живе за рахунок нагромаджених личинкою резервів). Зовні лялечка хоч і не схожа на імаго, проте вже має ряд ознак дорослої фази — зовнішні зачатки крил, ніг, вусиків, фасеткові очі тощо (рис. 155).
Часто перед залялькуванням личинка оточує себе коконом із шовку чи часточок субстрату, скріплених шовком або виділеннями мальпігієвих судин (у метеликів, пильщиків, їздців). Усередині цього кокона відбувається залялькування, кокон захищає лялечку від несприятливих впливів. Багато личинок жуків і гусені метеликів не роблять кокона, а заляльковуються в ґрунті, утворюючи порожнину — комірку, або колисочку, часто укріплену екскрементами. Іноді місцем залялькування є стебла рослин, скручене листя. Буває й відкрите залялькування, наприклад у денних метеликів.
Рис. 155. Типи лялечок:
а - відкрита (їздця); б - покрита (метелика); в - прихована (мухи)
Розрізняють три типи лялечок: відкриті, покриті й приховані (рис. 155). У відкритих зачатки крил і кінцівок вільні, лише притиснуті до тіла (більшість жуків, перетинчастокрилих, бліх, багатьох двокрилих та ін.). У покритих лялечок імагінальні придатки тісно зрощені з тілом унаслідок того, що личинка при останньому линянні виділяє секрет, який при затвердінні вкриває лялечку твердою оболонкою (більшість метеликів, деякі жуки та ін.). Приховані лялечки вкриті шкіркою останньої личинкової стадії, яка не скидається, а утворює несправжній кокон — пупарій, усередині якого міститься типова вільна лялечка (у вищих двокрилих, або мух).
Лялечка нерухома, але це тільки зовнішнє враження: на цій фазі відбуваються інтенсивні процеси внутрішньої перебудови —гістоліз та гістогенез. Оскільки характер руху імаго, особливо політ, і його живлення інакші, ніж у личинкової фази, то і м’язи, і травна система, а часто й ротовий апарат, кінцівки та інші личинкові органи мають зовсім перебудуватися. Більшість органів личинки руйнується; цей процес зветься гістолізом. Він відбувається фагоцитозом, автолізом (самоперетравлення), а також за допомогою ферментів. У різних комах переважає той чи інший механізм, але найчастіше діють усі три: гемоцити виділяють у гемолімфу ферменти, які руйнують тканини; їх залишки фагоцитукуються гемоцитами. Крім того, в деяких тканинах відбувається автоліз.
Коли гістоліз досягає свого піку, внутрішні органи лялечки перетворюються на напіврідку масу, яка складається з гемолімфи, збагаченої продуктами розпаду. Не руйнуються лише нервова, статева системи та спинна кровоносна судина. Нервова система може доповнюватися новими клітинами, в ній можуть концентруватися ганглії, проте вона ніколи не втрачає своєї цілісності.
Паралельно з гістолізом відбувається гістогенез — побудова імагінальних органів. Органи дорослої комахи формуються з особливих зачатків — імагінальних дисків (рис. 156). Це невеличкі скупчення недиференційованих клітин, розташовані в певних місцях тіла личинки. Кожен імагінальний диск має своє призначення: є диски, з яких утворюються крила, кінцівки, певні ділянки кишечника, фасеткові очі тощо. Імагінальні диски закладаються ще під час ембріонального розвитку або в ранніх личинок; протягом личинкового розвитку вони ростуть, але не диференціюються. Тільки в личинки останньої стадії перед залялькуванням їхні клітини диференціюються; в лялечки вони розгортаються, утворюючи імагінальні органи.
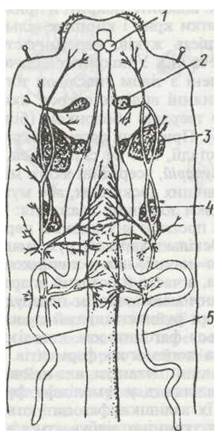
Рис. 156. Імагінальні диски гусені капустяного білана:
1 - мозок; 2 - передньогрудний зачаток; 3 - зачатки передніх крил; 4 - зачатки задніх крил; 5 - шовковидільна залоза; 6 - кишечник
Важливу роль у метаморфозі відіграє жирове тіло. На личинковій фазі в ньому нагромаджуються поживні речовини (жири, білки, вуглеводи), що використовуються як пластичний матеріал і джерело енергії при побудові імагінальних органів.
Метаморфоз комах відбувається під контролем гормональної (ендокринної) системи (рис. 157). Як уже зазначалося, метаморфоз регулює нейроендокринний комплекс органів, до якого входять нейросекреторні клітини мозку, кардіальні тіла, прилеглі тіла та парна перед- ньогрудна, або проторакаль- на, залоза. Нейросекреторні клітини мозку виробляють активаційний гормон. Через довгі аксони цих клітин він надходить до кардіальних тіл, а з них —у гемолімфу. У личинок активаційний гормон стимулює роботу проторакальних залоз, які починають виділяти линяльний гормон — екдізон. Останній впливає на клітини гіподерми, спричиняючи спочатку синтез ферментів, які розчиняють стару кутикулу, а пізніше — синтез матеріалу для побудови нової кутикули. Крім того, екдізон стимулює ріст і диференціацію всіх тканин личинки, особливо в гонадах, імагінальних дисках тощо.
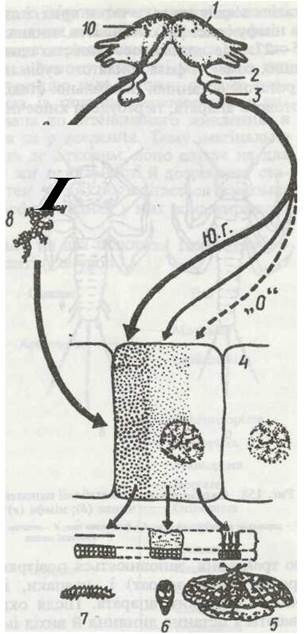
Рис. 157. Схема ендокринної системи, що бере участь у регуляції метаморфозу комах:
1 — головний мозок; 2 — кардіальні тіла; 3 — прилеглі тіла; 4 — гіподермальні клітини; 5 — імаго; 6 — лялечка; 7 — личинка; 8 — проторакальна залоза; 9 — мозковий гормон; 10 — нейросекреторні клітини; ю.г, —ювенільний гормон; «о» - відсутність гормону
Паралельно з екдізоном при личинкових линяннях виділяється ювенільний (юнацький) гормон, що гальмує метаморфоз, тобто перешкоджає линянню личинки на дорослу фазу. Наявність ювенільного гормону в гемолімфі визначає характер линяння. При високій концентрації ювенільного гормону екдізон гальмує розвиток імагінальних органів та стимулює секрецію личинкової кутикули, і линяння завершується появою чергової личинкової стадії. Зниження концентрації ювенільного гормону визначає линяння личинки на лялечку (або останню личинкову стадію при неповному метаморфозі). Припинення надходження ювенільного гормону в гемолімфу обумовлює линяння лялечки (або останньої личинкової стадії при неповному перетворенні) на імаго.
Походження різних типів метаморфозу становить предмет дискусій. Одні дослідники вважають, що повне перетворення походить від неповного, інші стверджують незалежне походження різних типів перетворення. Найбільш імовірно, що повне та неповне перетворення незалежно походять від архіметаболії — метаморфозу нижчих комах, у яких є личинкові та німфальні стадії. Серед сучасних комах архіметаболія відома у щетинкохвісток (підклас Apterygota) та одноденок (ряд Ерhemeroptera). В останніх з яйця виходить передличинка, або «личиночка», яка ще оточена ембріональною кутикулою, має майже гомономну метамерію, недорозвинені (з неповним числом члеників) антени й хвостові нитки, п'ять простих вічок і живиться ембріональним жовтком. Після першого линяння личиночка переходить в личинку, яка має вже цілком сформовані антени й хвостові нитки, фасеткові очі, зачатки трахейних зябер. На 5—6-й стадіях з'являються зачатки крил, і личинка перетворюється на німфу (рис. 158). Кількість линянь в одноденок становить 25—27. Ще однією особливістю одноденок, на відміну від інших комах, є фаза крилатого субімаго, здатного до польоту. Протягом останньої німфальної стадії починається редукція ротового апарата, перебудова кишечника (він втрачає функцію травлення, заповнюється повітрям і перетворюється на аеростатичний апарат) і, навпаки, інтенсивний розвиток крил та статевого апарата. Після окрилення субімаго відбуваються останнє линяння й вихід імаго. Схематично життєвий цикл одноденок можна показати так:
Рис. 158. Окремі фази архіметаболії одноденок: «личиночка» (а); личинка (б); німфа (в):
Ембріонізація личинкового періоду розвитку (тобто проходження цих стадій в яйці) призводить до неповного перетворення. Навпаки, конденсація німфального періоду у фазу лялечки (замість поступового перетворення) при збереженні личинкового періоду, приводить до повного перетворення.
Виникнення повного перетворення пояснюється тим, що личинка, як правило, живе в іншому субстраті, ніж імаго, що виключає трофічну конкуренцію між різними фазами розвитку. Крім того, у комах із повним перетворенням личинкова фаза спеціалізована до інтенсивного живлення, а імаго — до розмноження та розселення. Тому імагінальне живлення часто називають додатковим: воно слугує не для росту, а для підтримання життєдіяльності й дозрівання статевих продуктів. Подібна тенденція спостерігається й у комах з іншими типами метаморфозу, однак у них вона виражена значно слабше.
Клас Insecta поділяється на два підкласи: Первиннобезкрилі (Apterygota) та Крилаті (Pterygota).