ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 2 - Г.Й. Щербак - 1996
ТИП ЧЛЕНИСТОНОГІ (ARTHROFODA)
ПІДТИП ХЕЛІЦЕРОВІ (СНЕLIСЕRАТА)
КЛАС ПАВУКОПОДІБНІ (ARACHNIDA)
Павукоподібні поширені по всій земній кулі, основна маса видів — вільноживучі наземні тварини, й лише серед кліщів є паразити рослин і тварин, а також мешканці солоних і прісних вод. Описано близько 60 тис. видів.
Павукоподібним властиві всі ознаки хеліцерових, зокрема, поділ тіла на головогруди, що несуть хеліцери, педипальпи й чотири пари ходильних ніг, та черевце, на якому зрідка є видозмінені кінцівки (рис. 189).

Рис. 189. Павукоподібні:
й - скорпіон Euscorpius carpaticus; б - псевдоскорпіон Chelifer cancroides; в - сольпуга Galeodcs aianeoides; г - павук Latrodectus tredecimguttatus; д - кененія Kenenia mirabilis; e - кліпі Ixodes ricinus;
Сегменти головогрудей у більшості павукоподібних злиті в суцільну масу й укриті єдиним спинним головогрудним щитом. Тільки в сольпуг, а також тартарид та кененій (невеликі групи павукоподібних, перша поширена в екваторіальному поясі, друга — у південних районах з високою вологістю) в одну тагму злиті лише чотири передні сегменти головогрудей, а задні два сегменти вільні, й кожен укритий власним тергітом. Сегментація черевця у різних груп павукоподібних значно відрізняється, і його будову ми розглянемо конкретно при вивченні кожної окремої групи.
Перша пара кінцівок — хеліцери — найчастіше закінчуються клішнею, значно рідше вони гачкоподібні (наприклад, у павуків). Хеліцери, як звичайно, дво-, тричленикові; кінцевий членик відіграє роль рухомого пальця клішні або гачка. У скорпіонів та псевдоскорпіонів педипальпи закінчуються також потужною клішнею, а в сольпуг стають цілком схожими на ходильні ноги. Наступні чотири пари головогрудних кінцівок мають по шість-сім члеників і виконують роль ходильних ніг.
Зовні тіло павукоподібних укрите багатошаровою кутикулою, під якою залягає шар плескатих клітин гіподерми. Тонка будова кутикули в різних груп павукоподібних має відмінності. Ступінь склеротизації покривів різна у різних груп і видів, проте, як правило, особливо твердими є окремі ділянки тіла — щитки та членики ніг, а ділянки між ними тонші, еластичні.
Похідними гіподермального епітелію є численні залози: пахучі (у косариків), павутинні (у павуків), отруйні (у скорпіонів і павуків) тощо.
У павукоподібних, особливо у скорпіонів, є добре розвинений внутрішній скелет, подібний до такого у мечохвостів.
М'язова система дуже складна. Наприклад, у тілі скорпіона налічується не менше ніж 150 (частково парних) м'язів, не враховуючи м'язів кінцівок.
Травна система в павукоподібних, як і в інших членистоногих, складається з трьох відділів, проте задній (екгодер- мальний) відділ кишечника у них набагато коротший.
Будова травної системи у представників різних груп дещо відмінна (рис. 190), особливо задньої частини ентодермаль- ного відділу кишечника. В усіх павукоподібних є мускуляста глотка, що функціонує, здебільшого, за принципом насоса, через який усмоктується напіврідка їжа. Глотка переходить у тонкий стравохід, у якому в деяких павуків є ще одне розширення — смоктальний шлунок. Далі йде середня кишка, її передня частина часто розширюється (іноді стає мішкоподібною), утворюючи шлунок, від якого відходять бічні випини (дивертикули) різної довжини та товщини, завдяки чому значно збільшуються об'єм та поверхня стінок, де відбуваються травлення та всмоктування їжі. В середню кишку більшості павукоподібних відкриваються протоки масивної парної залози (печінки), функції якої відповідають сукупності функцій печінки та підшлункової залози хребетних. У печінці частково або в основному перетравлюється їжа (порожнинне та внутрішньоклітинне травлення), в її клітинах нагромаджуються поживні речовини. У скорпіонів, крім печінки, в головогрудях є ще шлункова залоза, яка складається з численних залозистих часток і відкривається в шлунок двома протоками. Гістологічно й функціонально вона не відрізняється від печінки. У павукоподібних дуже поширене вїгутршшьоклітинне травлення, що здійснюється клітинами як середньої кишки, так і печінки. Задня ектодермальна кишка має вигляд короткої прямої трубки.
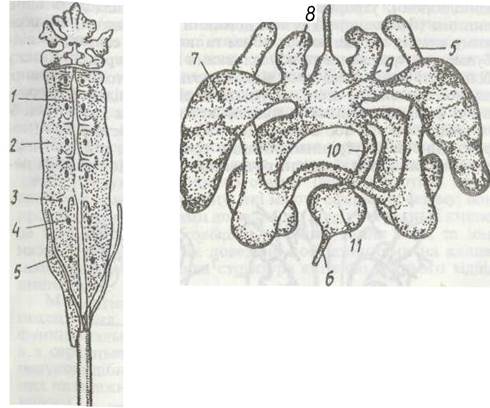
Рис. 190. Схема будови травної системи павукоподібних: а - скорпіоне; б - кліща Yarroa jacobsoni
Для павукоподібних характерне також позакишкове травлення. Прокушуючи покриви здобичі, вони виливають у неї травний сік, до складу якого входять сильнодіючі травні ферменти, що утворюються в середній кишці й частково в слинних залозах (якщо вони є). Розщеплюючи білки, вони перетворюють тіло жертви на напіврідку масу, придатну до всмоктування. Як уже згадувалося, більшість груп павукоподібних — хижаки, рідше вони поїдають різні рослинні рештки (косарики, деякі кліщі), ссуть соки рослин або живляться кров'ю хребетних тварин, у тому числі й людини (кліщі).
Основними органами виділення павукоподібних є мальпігієві судини — сліпо замкнені з одного кінця, іноді розгалужені трубки, які відкриваються в середню кишку в задній її частині (рис. 191). Вони функціонують у комплексі із заднім відділом середньої кишки, де часто утворюються спеціалізовані розширені ділянки (клоакальна сумка у павуків, ректальний міхур у частини кліщів). Гістологічна структура цих ділянок мало відрізняється від такої мальпііієвих судин, які в них впадають. Основним продуктом виділення в павукоподібних є гуанін. Це нерозчинна у воді речовина, яка подібно до сечової кислоти комах дає змогу павукоподібним заощаджувати в організмі воду. У мальпігієвих судинах розчинні продукти азотистого обміну перетворюються на нерозчинні кристали гуаніну, які мають сферичну форму; вони просуваються судинами до заднього відділу середньої кишки, де відбувається реабсорбція — всмоктування води та іонів назад у гемолімфу. Як доведено дослідженням и на кліщах, цьому сприяє особлива структура епітелію заднього відділу кишечника.
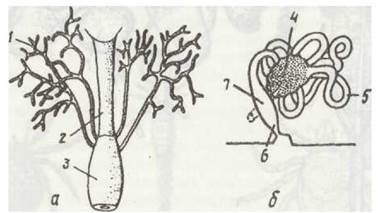
Рис. 191. Органи виділення павукоподібних:
а - мальпігієві судини Araneus diadematus; б - коксзльна залоза скорпіона
Мальпігієві судини павукоподібних мають ентодермальне походження, на відміну від аналогічних утворень комах, і функціонально пов'язані не із задньою кишкою, як у комах, а з середньою. Це свідчить, що мальпігієві судини комах і павукоподібних не є гомологічними утворами, а виникли у них незалежно як пристосування до життя в умовах дефіциту волога.
Крім мальпігієвих судин, функцію виділення виконують парні коксальні залози, що мають у деталях різну будову, проте в більшості випадків складаються з кінцевого мішка, звивистого каналу (лабіринта) та більш-менш прямої вивідної протоки із сечовим міхурцем. Протока відкривається біля тазиків третьої-п'ятої пар кінцівок головогрудей. Коксальні залози розвинені добре у ембріонів і молодих особин, але найчастіше вони більш-менш атрофуються, й тільки в косариків зберігають видільну функцію протягом усього життя. У кененій, як вважають, коксальні залози є єдиними органами виділення.
У виділенні беруть участь також різні ділянки кишечника, навіть у форм, що мають добре розвинені спеціальні екскреторні органи. Особливо інтенсивно нагромаджуються продукти розпаду в епітелії дивертикул (павуки, косарики, псевдоскорпіони та інші) та задній частині ентодермальної середньої кишки. У частини тромбідіформних кліщів цей від діл повністю виконує функцію виділення й навіть не з'єднується з передньою частиною середньої кишки, яка сліпо замкнена.
Гуанін концентрується в клітинах кишечника, а потім викидається в його порожнину, або навіть уся клітина злущується і виводиться назовні. Крім того, в багатьох павукоподібних гуанін часто відкладається в тканинах і не виводиться зовсім.
У виділенні беруть участь також спеціальні, досить великі за розміром клітини — нефроцити, розташовані в порожнинах між органами; вони нагромаджують продукти виділення.
Органами дихання у павукоподібних є легеневі мішки (скорпіони, павуки), трахеї (сольпуги, сінокоси, псевдоскорпіони, кліщі) або ті й інші разом ( павуки). Кожен легеневий мішок удається всередину тіла від щілиноподібного отвору — стигми. У порожнину мішка вдаються численні паралельні одна одній листкоподібні пластинки — тонкі сплющені складки стінок легені, складені подібно до листків книги (рис. 192, а). Стінка кожної легеневої пластинки вкрита дуже тонкою кутикулою. На верхній стороні кожна пластинка має маленькі кутикулярні стовпчики, які не дають сусіднім пластинкам злипатися, завдяки чому в проміжках між пластинками завжди є повітря. Під кутикулою пластинок лежить шар гіподерми, а всередині — вузькі лакунарні порожнини, заповнені гемолімфою. Газообмін відбувається через тонкі стінки пластинок. Скорпіони мають чотири пари легеневих мішків, більшість павуків — одну, зрідка дві пари.
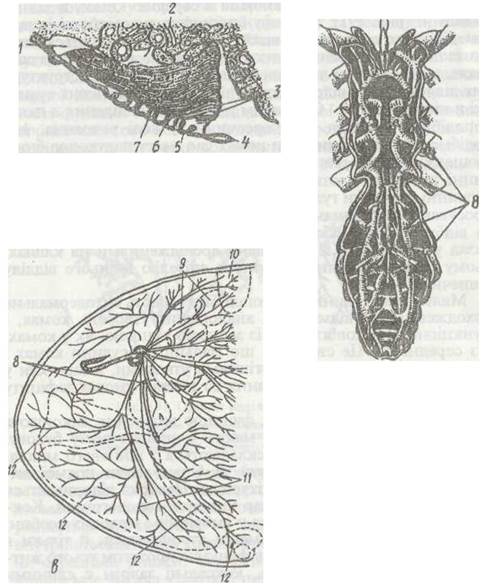
Рис. 192. Органи дихання павукоподібних:
а - зріз через легеневий кішок павука-хрестовика; б - головні трахейні стовбури сольпуги; в - схема будови трахейної системи кліща Vairoa jacobsoni;
Трахеї, як і в інших членистоногих, становлять систему різною мірою розгалужених трубок, що починаються спеціальними дихальними отворами (стигмами). Форма, кількість і місце розташування стигм, товщина трахейних стовбурів і ступінь їх розгалуження в кожній групі мають свої особливості (рис. 192, б, в). Найкраще розвинена трахейна система в сольпуг. Основні трахейні стовбури відкриваються в них кількома парами дихалець на головогрудях і черевці та одним непарним — на IV сегменті черевця. Трахеї, що відходять від дихалець, об'єднуються в могутні поздовжні стовбури, які зв'язані перемичками й посилають численні розгалуження до всіх органів. У стінках трахей сольпуг є спіральні потовщення кутикули, подібні до тенідіїв комах.
Більшість павуків, поряд із легенями, має ще й трахеї. Найчастіше у них є лише одне черевне дихальце, від якого відходять дві пари сліпо замкнених нерозгалужених трубочок, вистелених тоненькою кутикулою без спіральних потовщень. Трахеї омиваються гемолімфою.
У дрібних форм, зокрема в деяких кліщів і кененій, дихання відбувається через тонкі покриви тіла, і спеціальні органи дихання в них відсутні.
Ступінь розвитку кровоносної системи пов'язаний із розмірами тварин, почленованістю їхнього тіла та будовою органів дихання. З розвитком трахейної системи кровоносна система стає менш розвиненою.
Найкраще кровоносна система розвинена у скорпіонів. Серце в них має вигляд довгої трубки, що тягнеться на спинній стороні майже через усю передню частину черевця. Серце розташоване в тонкостінному перикардії й підвішене в ньому парними сполучнотканними тяжами (лігаментами). На спинній стороні серця є сім метамерно розташованих остій. Через них, завдяки спеціальним клапанам, гемолімфа проходить лише в одному напрямку — з перикардія в серце. Спереду і ззаду серце продовжується в передню та задню аорти, від яких відходять численні судини, що, розгалужуючись, несуть кров до всіх органів і тканин. Крім аорт, від серця відходять дев'ять пар бічних артерій; вісім із них занурюються в печінку, де утворюють численні розгалуження, що пронизують весь орган (рис, 193). Кінцеві гілочки всіх судин відкриті, й гемолімфа врешті- решт потрапляє в систему лакун та синусів, частина яких має власні сполучнотканинні стінки, їх можна назвати венами. У головогрудях та передчеревці з лакун венозна гемолімфа збирається в пару поздовжніх вентральних синусів, які біля легень утворюють розширення — легеневі синуси, котрі омивають легеневі мішки. Заходячи в лакуни легеневих пластинок, гемолімфа збагачується киснем. Із легеневих синусів сімома парами легеневих вен гемолімфа надходить до перикардія, а звідти через остії —до серця.
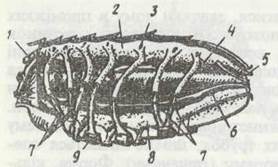
Рис. 193. Будова серця скорпіона:
1 — передня аорта; 2 - серце, вкрите перикардієм; 3 - ост; 4 -бічні артерії; 5 - задня аорта; 6 - черевний нервовий ланцюжок; 7 -легеневі вени; 8 - вентральний синус; 9 - легені
В інших павукоподібних серце відповідно до будови тіла вкорочується, кількість остій зменшується (наприклад, у павуків їх три-чотири, у косариків — одна-дві пари), залишається тільки передня аорта (павуки), зменшується кількість бічних судин (у павуків їх три пари), а в більшості кліщів відокремлена кровоносна система взагалі відсутня, й гемолімфа циркулює в порожнині тіла. Гемолімфа павукоподібних має дихальний пігмент гемоціанін.
Для багатьох павукоподібних характерний високий ступінь концентрації нервової системи, який прямо залежить від скорочення довжини їхнього тіла та злиття сегментів і тагм (рис. 194). Головний мозок складається з двох відділів: переднього — протоцеребрума, що іннервує очі, та заднього — тритоцеребрума, який посилає нерви до першої пари кінцівок (хеліцер).
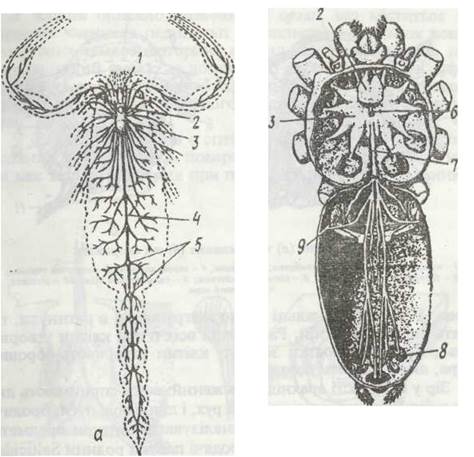
Рис. 194. Схема будови нервової системи скорпіона Androctonus (а) та павука Tegenaria (б):
1 - очі- 2 - мозок- 3 - підглотковий ганглій; 4 - черевний нервовий ланцюжок; 5 - ганглії на ньому; 6 - стравохід; 7 -черевний нервовий стовбур; 8 - нерви, що іннервують павутинні бородавки; 9 - легені
Найменш концентрована нервова система у скорпіонів (рис. 194, а). Вона складається з надглоткового й підглоткового гангліїв, що з'єднані короткими товстими конективами, та довгого черевного ланцюжка з сімома гангліями. Підглотковий ганглій іннервує 2—6 пар кінцівок. У павуків, сольпуг та деяких інших груп підглотковий ганглій і ганглії черевного ланцюжка зливаються в єдину гангліозну масу (рис. 194, б). У косариків, псевдоскорпіонів та кліщів усі ганглії зливаються в єдине гангліозне тіло (синганглій), розміщене навколо стравоходу.
Органи зору представлені простими очима, розташованими на верхній стороні головогрудей (рис. 195). Кількість їх у різних павукоподібних різна: у скорпіонів е пара серединних (медіальних) та дві—п'ять пар бічних; у павуків найчастіше вісім очей, розташованих у дві дуги; чотири з них лежать медіально, інші дві пари — по боках.
Бічні очі мають простішу будову (рис. 195, а). Це бокалоподібні очі, до складу яких входять опукла лінза, або кришталик, та сітківка, що складається з видовжених зорових (ретинальних) клітин. Останні згруповані по кілька, утворюючи численні ретинули з рабдомами, не відокремлені одна від одної пігментом. Базальні кінці зорових клітин зібрані в зоровий нерв. Гіподерма утворює навколо ока суцільне темне кільце.
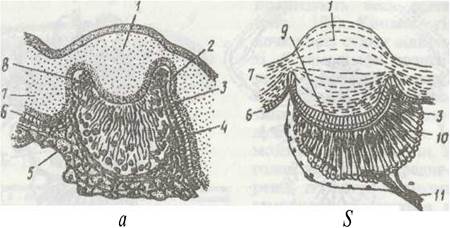
Рис. 195. Бічне (а) та серединне (б) очі скорпіона:
1 - кришталик; 2 - базальна мембрана; 3 - рабдом; 4 - зорові клітини; 5 - сполучна тканина; 6 - гіподерма; 7 - кутикула; 8 - пігментні клітини; 9 -склоподібне тіло; 10 —сітківка; 11 - зоровий нерв
Медіальні очі мають кришталик, склоподібне тіло й ретину, яка складається з ретинальних і пігментних клітин (рис. 195, 6). Ретинальні клітини згруповані в ретинули, по п'ять клітин у кожній. Рабдомери всіх п'яти клітин утворюють рабдом. Відростки зорових клітин утворюють зоровий нерв, який входить у мозок.
Зір у більшості арахнід обмежений: вони сприймають лише зміну інтенсивності світла й рух, і лише сольпуги, бродячі павуки та скорпіони здатні аналізувати контури предметів своїми медіальними очима. Бродячі павуки родини Salticidae відзначаються більш досконалим зором: спеціальні м'язи рухають очі, завдяки чому вони можуть стежити за здобиччю, залишаючися нерухомими.
На тілі та кінцівках є численні, різні за будовою та функціями, чутливі волоски. Поряд зі звичайними, трихощними сенсилами, у павукоподібних дуже поширені трихоботрії (рис, 196, а). Вони розташовані на педипальпах і ногах або на тілі (у частини кліщів). Довгий волосок, іноді потовщений на кінці, прикріплюється тонкою мембраною на дні лійкоподібного заглиблення в кутикулі; до його основи підходить група чутливих клітин. Найменше коливання повітря чи субстрату спричиняє його зміщення, яке сприймається чутливими клітинами.
Сенсили часто зібрані разом, утворюючи органи. Для арахнід характерні так звані ліроподібні органи, розташовані на тулубі та кінцівках. Це мікроскопічні щілини в кутикулі, затягнуті тонкою мембраною, до якої підходить відросток чутливої клітини (рис. 196, б, в). Вважають, що ці органи — механорецептори, які сприймають ступінь натягу кутикули.
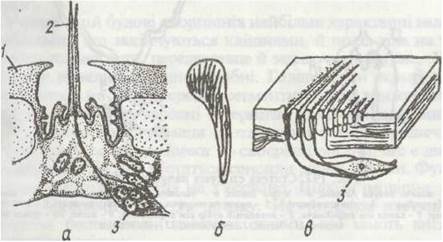
Рис. 196. Органи чуття павукоподібних:
а - трихобогрія; б, в -ліроподібний орган з поверхні та в розрізі відповідно; 1 -кутикула; 2 - волосок; 3 - чутлива нервова клітина
У кліщів описано пальпальний орган, що міститься на кінцевих члениках педипальп і складається з кількох конусоподібних хеморецепторних сенсил. За функціями це, передусім, орган смаку, за допомогою якого кровосисні кліщі вибирають на тілі тварини-годувальника місце для кровоссання. Крім того, частина його сенсил є нюховими рецепторами.
У іксодощних кліщів є специфічний орган Талера, розташований на дорзальній поверхні лапок передньої пари ніг; він має велике значення при пошуках кліщем годувальника.
Павукоподібні роздільностатеві. Статеві залози містяться в черевці й можуть бути парними або непарними. Яйцепроводи та сім'япроводи парні, але назовні відкриваються непарним статевим отвором. У самиць, як звичайно, є розширення яйцепроводу — матка та сім'яприймачі, в яких зберігається сперма. В деталях будова статевої системи у представників різних рядів різна (рис. 197). Запліднення або сперматофорне, або відбувається при копуляції. Більш детально це питання буде розглянуто далі.
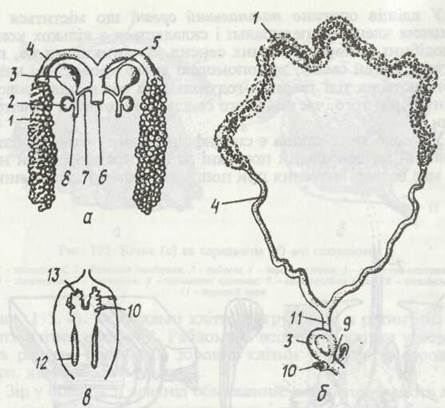
Рис. 197. Статева система павукоподібних:
а - жіноча павука Araneus; б - кліща Ixodes; в - чоловіча Araneus; 1- яєчник; 2 - залоза сім’яприймача; 3 - сім’яприймач; 4,5- парна та непарна частини яйцепроводу, 6 - статевий отвір; 7 - канал сім’яприймача; 8 - зовнішній отвір сім’яприймача; 9 - піхва; 10 - придаткові залози; 11 - матка; 12 -сім’яник; 13 - сім'япровід
Більшість арахнід відкладають яйця, хоч серед них є й живородні. Плодючість дуже різна — від кількох сотень яєць до 30 000.
Яйця більшості арахнід великі й багаті на жовток, тому дробіння в них, як і в інших членистоногих, найчастіше поверхневе, неповне. Зародок формується переважно за рахунок зародкової смужки. У зародка сегментація буває виражена краще, його тіло має більше сегментів, ніж доросла тварина. Наприклад, у зародків павуків черевце складається з 12 сегментів, як у скорпіонів, причому 4—5 передніх сегментів мають зачатки ніг. У зародків скорпіонів утворюються зачатки черевних ніжок. У павуків і скорпіонів можна простежити, як певні зачатки черевних ніг перетворюються на легеневі мішки.
Майже в усіх павукоподібних розвиток прямий, супроводжується ростом і дорозвиненням певних органів під час линянь.
Система класу павукоподібних перебуває в стадії розробки. Немає єдиної думки ні про кількість таксонів, що в ньому об'єднані, ні про їхній ранг. До того ж висловлюються думки про штучність класу в цілому. Ми розглянемо ряд груп павукоподібних, яких приймаємо в ранзі підкласу.