ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 3 - Г.Й. Щербак - 1997
ТИП ЩЕТИНКОЩЕЛЕПНІ, АБО МОРСЬКІ СТРІЛКИ (CHAETOGNATHA)
КЛАС ЩЕТИНКОЩЕЛЕПНІ, АБО МОРСЬКІ СТРІЛКИ (CHAETOGNATHA)
Щетинкощелепні —винятково морські хижаки, які погано переносять опріснення, лише поодинокі види пристосувались до пониженої солоності води. Описано близько 150 видів, з яких три види знайдено в Чорному морі ( Sagitta euxina — ендемік цього моря) і один — в Азовському. Основна маса видів живе в товщі води, на різних глибинах, перебуваючи в постійному русі. У деяких видів є пристосування для ширяння у воді —широкі крилоподібні плавці (рід Krohnitta), зменшення маси тіла за рахунок редукції зубчиків, щетинок, м’язової тканини, які поряд зі збільшенням об’єму тіла знижують його питому вагу (рід Flaccisagitta) тощо. Усього дев’ять видів родини Spadellidae живуть біля дна як на мілководді, де вони маневрують між водоростями та камінням, так і на більших глибинах (до 1000 м).
Розміри морських стрілок коливаються від 0,5 до 9 см. Найдрібніші види — придонні, найбільші — північні нектонні форми.
Як правило, щетинко- щелепні безбарвні, і тому їх важко помітити у воді. Червонуваті, оранжево- коричневі, жовто-зелені стрілки трапляються лише серед видів, що ведуть придонний спосіб життя.
Усе тіло щетинкощелепних вкрите тонкою кутикулою, під якою міститься одношаровий, так званий пластирний, епітелій (рис. 123). Особливість його будови полягає в тому, що краї клітин хвилеподібні й з’єднані так, що опуклість однієї клітини входить у заглиблення сусідньої. Таке зубчасте зчеплення надає епітелію розтяжність та гнучкість — якості, що необхідні при швидких дорзовентральних вигинах тіла під час полювання, оскільки вони перешкоджають розриву покривів.
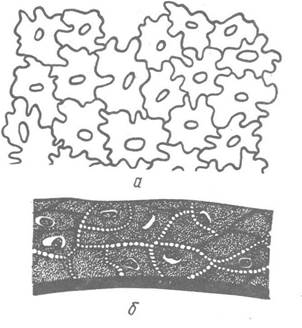
Рис. 123. Покриви щетинкощелепних: а — «пластирний епітелій»; б — багатошаровий епітелій у шийній ділянці
Окремі ділянки, зокрема між головним та тулубним відділами над мозком (комірцева зона), тулубної та хвостової частин тіла, вкриті, крім шару покривного епітелію, ще одним шаром більш пухких клітин, які часто розростаються в багаторядний шар. Функція цих клітин захисна, наприклад товстий шар епітелію в комірцевій зоні править за демпфер (глушник) для пом’якшення удару об здобич.
Похідними покривів є ряд структур, зокрема щетинки та зубці ловецького апарату, так званий капор та плавці. Щетинки та зубці мають схожу будову —це хітинізовані утвори, зовні вкриті кутикулою, всередині яких проходить канал, наповнений пульпою. Щетинки, як правило, довші й серпоподібно вигнуті, їх внутрішній край часто зубчастий; до основної частини щетинок прикріплені м’язи, що рухають їх під
час захоплення здобичі. Зубці, як правило, більш короткі і менш зігнуті, ніж щетинки. Щетинок завжди два ряди, зубців — здебільшого два ряди (рис. 124).
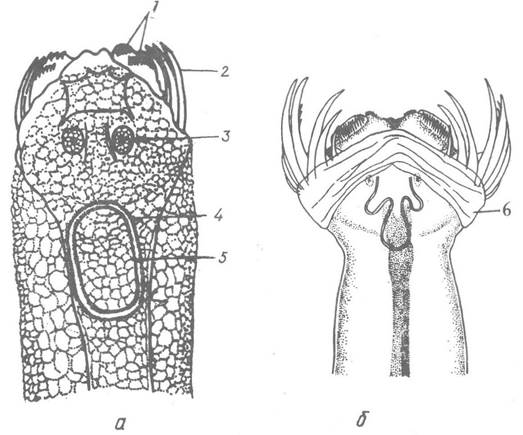
Рис. 124. Передній кінець тіла щетинкощелепних зі спинної сторони:
а — Pterosagitta draco; б — Flaccisagitta inflata (вигляд через покриви); 1 — передній та задній ряди зубців; 2 — щетинки; 3 — очі; 4,5 — зовнішнє та внутрішнє кільця миготливої петлі; б — капор
Капор — унікальний орган, який є тільки в морських стрілок. Це шкірна складка голови, яка може миттєво відкидатись, відкриваючи ловецький апарат, і так само швидко його закривати. При будь-яких переміщеннях капор закриває ловецький апарат, чим збільшується обтічність тіла, у момент захоплення здобичі капор різким рухом відкидається назад, а щетинки та зубчики пронизують жертву. Рух капора відбувається за допомогою двох антагоністичних м’язів.
По боках тіла розташовані одна або дві пари бічних плавців, які виконують переважно функцію керма, а на кінці тіла міститься хвостовий плавець — основний локомоторний орган. У променях плавців знайдено особливу речовину — еластощин, що характерний ще для круглоротих та риб.
Під покривним епітелієм розташована базальна мембрана — пружна пластинка, що зумовлює форму тіла; вона слугує антагоністом відносно поздовжньої мускулатури при згинанні тіла в дорзовентральному напрямі, а також його скелетом — до базальної мембрани прикріплюються різні групи м’язів.
М’язова система добре розвинена: у головному відділі знаходяться численні спеціалізовані пучки м’язів, які рухають капор, щетинки, зубці тощо. У тулубному та хвостовому відділах м’язи згруповані в чотири поздовжні тяжі, які різною мірою розвинені в нектонних, придонних та видів, що здатні до ширяння. Усі м’язи мають поперечно-смугасту будову.
У дорослих щетинкощелепних є об’ємна порожнина тіла, не вистелена перитонеальним епітелієм, хоча в процесі ембріонального розвитку закладається пара целомічних мішків, які швидко втрачають просвіт, а з клітин їх стінок походять м’язи та інші мезодермальні органи. Через втрату перитонеального епітелію морських стрілок часто відносять до тварин із первинною порожниною, але це питання ще остаточно не вирішене.
Порожнина в дорослих особин розділена двома мезодермальними перетинками на три відділи; крім того, в тулубному відділі є поздовжня перетинка, на якій підвішений кишечник.
Травна система починається ротовою лійкою — заглибленням перед ротовим отвором, у покривах якого є численні секреторні клітини, виділення яких використовується для змащення здобичі, що полегшує її заковтування. Ротовий отвір веде в розширену мускулясту глотку, яка потім звужується і переходить на рівні тулубно-головної перетинки в ентодермальну частину кишечника. На самому її початку в деяких видів є парні кишенеподібні вирости, які, ймовірно, захищають кишечник від розриву при заковтуванні великої за об’ємом здобичі. Перед тулубно-хвостовою перетинкою середня кишка переходить у коротку задню кишку, що відкривається анальним отвором далеко від заднього кінця тіла (рис. 125).
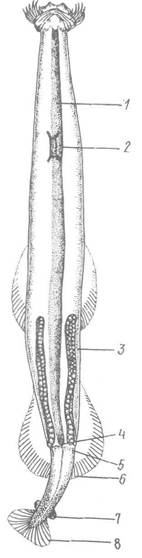
Рис. 125. Внутрішня будова щетинкощелепних:
1 — кишечник; 2 — черевний ганглій; 3 — яєчник; 4 — анус; 5 — сім’яник; 6 — тулубно-хвостова перетинка; 7 — сім’яний мішечок; 8 — хвостовий плавець
Полюють морські стрілки переважно вночі. Поїдають здебільшого рачків, але нападають і на личинок риб та власну молодь.
Іноді розмір здобичі значно перевищує розмір власного тіла стрілки, і тоді здобич заковтується поступово.
Видільної та кровоносної систем немає, газообмін відбувається через поверхню тіла.
Нервова система розвинена добре. Вона складається з непарного церебрального (головного) ганглію, або мозку,
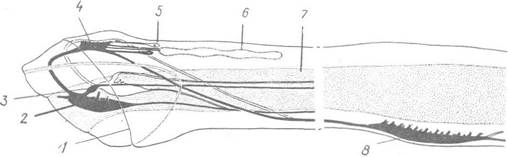
Рис. 126. Центральна нервова система Sagitta:
1 — край напівзрізаного капора; 2 — вестибулярний ганглій; 3 — глотковий ганглій; 4 — церебральний ганглій; 5 — око; 6 — миготлива петля; 7 — шлунок; 8 — черевний ганглій
розташованого на дорзальній стороні голови, пари вестибулярних гангліїв, розташованих поблизу мозку, пари дрібніших глоткових гангліїв і непарного великого черевного ганглію, який міститься в тулубному відділі (рис. 126); усі ганглії з’єднані конективами. Від гангліїв відходять нервові волокна до всіх органів.
Органи чуття представлені очима і так званою миготливою петлею. Очі розташовані на спинній стороні голови позаду мозку, вони складаються з п’яти пігментованих бокалів, розташованих у такий спосіб, що стрілки сприймають зображення не тільки зі спинної, але й з черевної та бічних сторін. Оскільки їх тіло прозоре, поле зору в них становить 360 °.
Миготлива петля має вигляд двох валків, що замкнені у вигляді зовнішнього та внутрішнього кілець. Петля розташована звичайно позаду очей (див. рис. 124) і тільки в небагатьох родів — перед ними. Зовнішній валок у всіх щетинко-щелегших вкритий вібруючими війками, тонка будова внутрішнього кільця різна в представників різних родів і в частини з них бере участь у процесі запліднення, про що йдеться далі. Основна функція миготливої петлі у всіх щетинкощелепних сенсорно-моторна, подібна до функцій бічної лінії в риб. Цим органом морські стрілки сприймають коливання води — сигнал, що йде від об’єктів живлення або особин того самого виду в період розмноження.
Щетинкощелепні — гермафродита і мають досить складну статеву систему. Жіночий статевий апарат міститься в задній частині тулубного відділу і складається з пари яєчників, розташованих по боках кишечника. До них прилягають сім’яні канальці, що виконують функцію сім’яприймачів; вони відкриваються назовні на бічних стінках тіла, біля статевих сосочків. Кінець канальця, що обернений до яєчника, замкнений сліпо; існує думка, що зв’язок з яєчником з’являється лише в період розмноження.
Чоловічий статевий апарат міститься в хвостовому відділі. До нього належать пара сім’яників, розташованих позаду тулубно-хвостової перетинки по боках тіла, та сім’яні мішечки, що виступають по боках тіла (див. рис. 125). Форма та місце їх знаходження варіюють у різних видів. Сперматозоїди на різних стадіях зрілості виходять у порожнину тіла, звідки після дозрівання по вивідній протоці надходять у сім’яні мішечки, де з’єднуються клейким секретом в сперматофор. Подальша доля сперматофора не однакова у представників різних груп, але у всіх щетинкощелепних запліднення внутрішнє. У нектонних форм (найкраще цей процес вивчено для стрілок родини Sagittidae) сперматофор (внаслідок різкого руху хвостового відділу) звільняється з сім’яного мішечка через розрив його стінки і потрапляє на хвостовий плавець, який обгортає сперматофор. Відсутність м’язів у плавцях наводить на думку про те, що обгортання відбувається через виділення з сперматофору клейкої речовини.
Обмін сперматофорами між двома стрілками відбувається при паруванні, коли вони розміщуються так, що хвостові плавці однієї особини торкаються задніх бічних плавців іншої. При цьому сперматофор однієї стрілки перекладається на бічний плавець іншої, після чого край цього плавця також загинається навколо нього. Звідси сперматозоїди потрапляють до сім’яприймачів.
У видів родини Spadellidae, що ведуть придонний спосіб життя (наприклад, у Spadella cephaloptera), у русі сперматозоїдів беруть участь клітини внутрішнього кільця миготливої петлі. Вони виділяють секрет, що надходить у кільцеву борозенку між зовнішнім та внутрішнім кільцями і далі стікає по середній лінії тіла до рівня тулубно-хвостової перетинки. Тут потік роздвоюється і прямує до отворів сім’я- приймачїв, де збирається у великій кількості, заходячи і в середину останніх. Під час копуляції дві особини з’єднуються між собою, як показано на рис. 127. Вони швидко рухаються подібно ножицям, і при цьому сперматофор однієї особини потрапляє на спинну сторону тулуба другої нижче миготливої петлі. Сперматофор руйнується в хвостовій частині, сперматозоїди виходять з нього і потрапляють на потік секрету, який несе їх до отворів сім’янриймачів, Якщо сперматофор потрапляє в інше місце тулуба, у тому числі і в середину петлі, він гине. У всіх щетинкощелепних можливе самозапліднення. Запліднені яйця активно проходять через епітелій яєчника і далі, прориваючи стінку тіла, виходять біля статевого сосочка назовні.
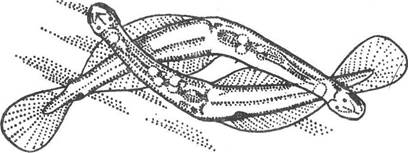
Рис. 127. Копуляція в Spadella cephaloptera
Яйця відкладаються по одному, або склеюються клейким секретом, що виділяється клітинами статевого сосочка в грона; в обох випадках вони прикріплюються до підводних предметів за допомогою того самого клейкого секрету. У холодноводних видів родини Eukrohniidae розвиток яєць відбувається в спеціальній сумці.
Дробіння яєць повне, рівномірне і починається як спіральне, потім втрачає правильність. Гаструляція проходить шляхом інвагінації. Бластопор у гаструли замикається, і значно пізніше на цьому місці виникає анус, а вторинний рот утворюється на протилежному (передньому) кінці зародка (рис. 128). Целом утворюється ентероцельно, шляхом вростання в ентодермальний зачаток первинного кишечника двох поздовжніх перетинок, які поділяють його порожнину на центральну частину — ентодермальну (власне кишечник) та дві бічні — мезодермальні з порожнинами всередині кожної (целоми). Згодом целом заростає, а клітинний матеріал його стінок (мезодерма) використовується на побудову мускулатури й інших похідних мезодерми.
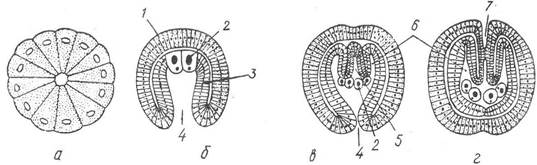
Рис. 128. Ембріональний розвиток щетинкощелепних:
а — бластула; б — гаструла; в—г — закладка целома та вторинного рота; 1 — ектодерма; 2 — первинні статеві клітини; 3 — ентодерма; 4 — бластопор; 5 — первинна кишка; 6 — бічні випинання первинної кишки (зачаток целома); 7 — вторинний рот
Через два дні після початку розвитку з яєць виходять молоді особини, схожі на дорослих. Вони живляться виключно дрібною їжею — бактеріями, одноклітинними водоростями тощо, поки в них не з’являться щетинки та зубчики ловецького апарату.
Морські стрілки відіграють суттєву роль у харчових ланцюгах морів та океанів. У період масового розмноження вони можуть бути конкурентами молоді планктон оїдних зграйних риб, проте дорослі щетинкощелепні в певні періоди становлять домінуючу їжу таких цінних промислових риб, як лососеві, оселедцеві та тріскові.
Щетинкощелепні вражають поєднанням у собі просто збудованого тіла та високоспеціалізованих тканин. У них немає дихальної, кровоносної і видільної систем та статевих проток; травна система у них представлена прямою кишкою без будь-яких додаткових залоз. Водночас стрілки мають поперечно-смугасту мускулатуру, подібну до мускулатури комах, багатошаровий покривний епітелій, схожий на такий у хребетних. В їхніх плавцях є еластоїдин, властивий рибам та круглоротим; вони мають високорозвинену нервову систему.
Морські стрілки досить давня група — їх відбитки відомі з нижнього кембрію.