ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 3 - Г.Й. Щербак - 1997
ТИП МОЛЮСКИ, АБО М’ЯКУНИ (MOLLUSCA)
КЛАС ДВОСТУЛКОВІ (BIVALVIA)
Двостулкові — виключно водяні тварини, що мешкають у морях, океанах та прісних водоймах. Розміри їх тіла — від кількох міліметрів до 1,5 метра (Tridacnagigas). Вони ведуть придонний спосіб життя і трапляються на різних глибинах. Більшість із них —повільно повзаючі форми, деякі нерухомо прикріплюються до субстрату. До цього класу належить близько 20000 видів. У прісних водоймах України їх налічують близько 150 видів, у Чорному та Азовському морях — понад 100 видів.
Двостулкові — це білатеральносиметричні молюски з двостулковою черепашкою, яка вкриває тіло з боків. Тіло їх здебільшого видовжене, більш-менш сплющене з боків і складається з тулуба, що заповнює верхню частину черепашки, і ноги, яка міститься на черевній стороні. Характерна особливість двостулкових —редукція голови. На передньому кінці тулуба розташований рот, на задньому — анальний отвір (рис. 25).
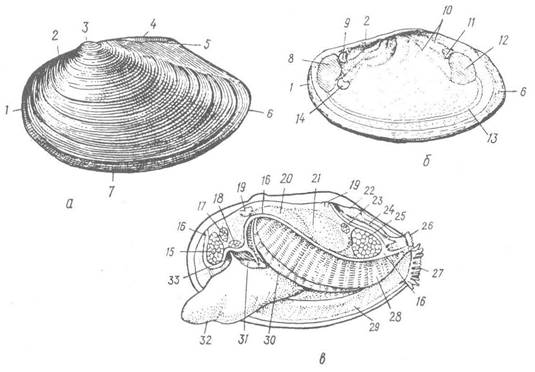
Рис. 25. Зовнішня будова беззубки (Anodonta cygnea):
а — зовнішній вигляд черепашки; б — внутрішня поверхня правої стулки; в — вигляд тіла молюска зліва, черепашку видалено; 1 — передній край черепашки; 2, 5 — спинний (замковий край); 3 — маківка; 4 — зовнішній лігамент; 6 — задній край; 7 — черевний край; 8 — відбиток переднього м’яза-замикача; 9 — відбиток переднього ретрактора ноги; 10 — сліди росту та переміщення м’язів; 11 — відбиток заднього ретрактора ноги; 12 — відбиток заднього м’яза-замикача; 13 —мантійна лінія; 14 — протрактор ноги; 15 —передній м’яз-замикач; 16 — лінія, по якій відрізано лівий мантійний листок; 17 — передній ретрактор ноги; 18— протрактор ноги; 19 — леватор (м’яз, що піднімає) ноги; 20 — кеберів орган; 21 — перикардій; 22 — спинний мантійний отвір; 23 — задній ретрактор ноги; 24 — спинний мантійний канал; 25 — задній м’яз-замикач; 26 — вивідний (анальний) сифон; 27 — ввідний (дихальний) сифон; 28 — ліва зовнішня напівзябра; 29 — мантія; 30 — ліва внутрішня напівзябра; 31 — ротова лопать; 32 — нога; 33 — рот
Ліва і права стулки черепашки рухомо з’єднані на спинній стороні еластичною зв’язкою — лігаментом, утвореним органічною речовиною — конхіоліном. Завдяки еластичності лігамента стулки при розслабленні м’язів-замикачів автоматично відкриваються. Крім того, у більшості двостулкових обидві стулки з’єднані між собою за допомогою замка, що складається із зубоподібних відростків внутрішньої поверхні спинного краю однієї стулки, які входять у заглибини протилежної. Деякі види, наприклад прісноводна беззубка (Anodonta), не мають замка, і стулки з’єднуються лише за допомогою лігамента.
Черепашки двостулкових мають різноманітну форму. У більшості обидві стулки однакові, проте є види, в яких вони різні. В устриці, наприклад, ліва стулка, якою вона прикріплюється до субстрату, значно більша й опукліша — у ній міститься все тіло м’якуна, тоді як права є лише покришкою. У деяких форм, наприклад у тередо деревоточця, або «корабельного черва» (Teredo) черепашка редукована: вона прикриває лише 1/20 довжини тіла (див. далі).
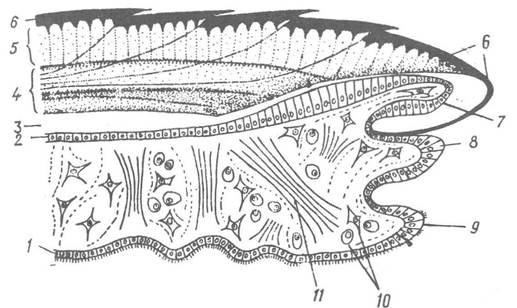
Рис. 26. Схема будови черепашки та мантії двостулкових молюсків:
1, 2 — внутрішній війчастий та зовнішній епітелій мантії; 3 — екстрапаліальна порожнина; 4, 5, 6 — перламутровий, призматичний та роговий (периостракум) шари черепашки; 7, 8, 9 — зовнішня, середня та внутрішня лопаті мантії; 10 — клітини сполучної тканини; 11 — м’язи сполучнотканинного шару мантії
Закривається черепашка м'язами-замикачами, яких буває два або один (за рахунок злиття двох м’язів або редукції одного з них). Вони мають вигляд товстих мускульних пучків, що йдуть упоперек тіла від однієї стулки до іншої (рис. 25, б, в). У місцях прикріплення м’язів до стулок на них утворюються відбитки, за якими можна зробити висновок про кількість та розташування м’язів-замикачів.
Під черепашкою розташована мантія, яка звішується з боків тіла у вигляді двох великих мантійних складок. Між мантією та черепашкою є дуже вузька екстрапаліальна порожнина (рис. 26), а між мантією і тілом залишається досить велика мантійна порожнина, в якій міститься мантійний комплекс органів: нога, зябра, осфрадії, ротові лопаті, ротовий, анальний, парні видільні та статеві отвори. На спинній стороні тіла обидві складки мантії зростаються. Коротенькими мантійними м’язами вільний край мантії прикріплюється до нижнього краю стулки черепашки, утворюючи на її внутрішній поверхні більш-менш глибокий слід — мантійну лінію (див. рис. 25, б). Край мантії, розташований нижче лінії прикріплення мантійних м’язів, потовщений і поділений вздовж на три лопаті — зовнішню, середню та внутрішню.
Краї мантійних складок у деяких місцях зростаються, залишаючи кілька отворів (2— 4), через які мантійна порожнина сполучається з оточуючим середовищем. Через такі отвори відбувається висування ноги, надходження води та їжі, вихід води, екскрементів, екскретів та статевих продуктів. Ввідний та вивідний отвори розташовані здебільшого на кінцях сифонів — порожнистих трубчастих виростів мантії (див. рис. 25, в). Сифони добре розвинені у форм, що зариваються в мул або пісок, у деяких видів їх довжина навіть перевищує довжину тіла (рис. 27, 28). Сифони можуть висуватися назовні і втягуватися всередину черепашки за допомогою особливих м’язів. У прісноводних перлівницевих (родина Unionidae) сифони короткі, а в кулькових (родина Sphaeriidae) їхня довжина може становити 0,3— 0,5 довжини тіла. По краю манти можуть розташовуватись щупальця, очі, залози тощо.
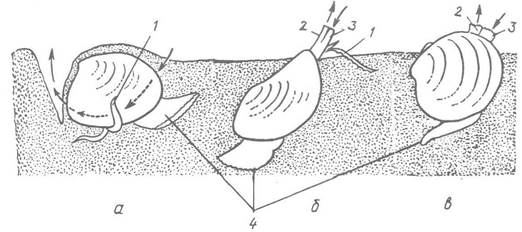
Рис. 27. Двостулкові молюски, які мешкають на поверхні дна та неглибоко закопуються в фунт:
а, б — горіховидка-нукула та куспідарія (надряд Protobranchia); в — циприна (надряд Autobranchia); 1 — ротові лопаті та придатки; 2 — вивідний сифон; 3 — ввідний сифон; 4 — нога. Стрілками позначено рух води
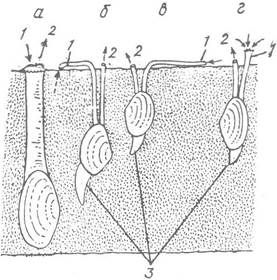
Рис. 28. Двостулкові молюски, які глибоко закопуються в ґрунт:
а — мія; б — скробікулярія; в — теліна; г — донакс (навряд Autobranchia);
1 — ввідний сифон; 2 — вивідний сифон; 3 — нога. Стрілками позначено рух води
Мантія —це багатошарове утворення (див. рис. 26). Зовні вона вкрита мантійним епітелієм, під яким міститься базальна мембрана, а під нею — потужний шар сполучної тканини (паренхіми), яка пронизана тонкими м’язовими волоконцями. Клітини сполучної тканини лежать пухко, між ними залишаються великі міжклітинні простори, а також лакуни, заповнені гемолімфою. Внутрішня поверхня мантії вистелена війчастим епітелієм. У сполучній тканині, поблизу від епітеліальних шарів, залягають численні залозисті клітини.
Встановлено, що всі шари черепашки утворюються виділеннями залозистого епітелію зовнішньої лопаті мантії, причому конхіоліновий шар виділяється її внутрішньою поверхнею, а призматичний — зовнішньою. На решті зовнішньої поверхні всієї мантії виділяються речовини, що використовуються на побудову перламутрового шару черепашки.
Епітелій мантії відіграє важливу роль в адсорбції йонів кальцію із зовнішнього середовища шляхом піноцитозу. Крім того, кальцій переноситься до мантії гемолімфою з органів травлення. Застосуванням міченого Са45 було встановлено, що слиз, який суцільним шаром вкриває зябра, інтенсивно адсорбує кальцій із води. Шматочки слизу, відриваючись від зябер, разом з їжею потрапляють до органів травлення, де кальцій звільнюється і переноситься гемолімфою до мантії. Секрет залозистого епітелію мантії виділяється в екстрапаліальну порожнину. Він містить органічні речовини, що входять до складу матрикса (органічної основи) черепашки, а також значну кількість кальцію, який відкладається на цьому матриксі у вигляді СаСО3.
Черепашка росте разом з твариною протягом усього її життя, причому вона наростає по вільному краю. Верхівки стулок є найстарішими частинами черепашки, до яких у процесі росту додаються все нові й нові ділянки. Завдяки цьому можна відрізнити щорічний приріст та визначити вік тварин, якщо умови росту в різні сезони року різні. Узимку вповільнення росту помітне на черепашці у вигляді згущення ліній наростання, по яких і визначають вік тварин.
Із секреторною діяльністю мантійного епітелію деяких двостулкових пов’язане утворення перлин. Формування перлів — це захисна реакція організму на потрапляння в нього стороннього тіла: піщинки, уламку черепашки, частинок органічної речовини, паразита. Клітини мантії починають огортати чужорідне тіло зовнішнім шаром мантії з утворенням навколо нього епітеліального, так званого перлинного,
мішечка (рис. 29). Епітелій мішечка продовжує нормально функціонувати, виділяючи всередину спочатку трохи периостракума, потім призматичний шар, і кінець-кінцем —перламутровий шар, тобто в тій самій послідовності, що й при рості черепашки. Так утворюється вільна перлина. Найцінніші перлини здатні утворювати морські перлівниці (Pinctada margaritifera) та європейська річкова перлова скойка (Margaritifera margaritifera).
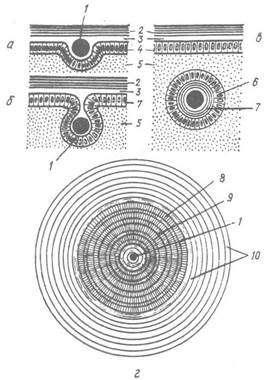
Рис. 29. Схема утворення перлини:
а — в — послідовні стадії; г — шліф через перлину; 1 — чужорідне тіло — «ядро» перлини; 2 — перламутровий шар черепашки; 3 — екстрапаліальна порожнина; 4 — зовнішній епітелій мантії; 5 — сполучна тканина мантії; 6 — мантійний мішечок; 7 — перлина; 8 — призматичні шари; 9 — шари конхіоліну; 10 — шари перламутру
У мантійній порожнині двостулкових міститься орган руху — нога. Залежно від способу життя двостулкових нога може мати різну форму. У деяких найпримітивніших представників (надряд Ргоtobranchia) нога має плоску повзаючу підошву. Здебільшого ж вона клиноподібної форми, дуже сплюснута з боків і загострена на кінці (див. рис. 27, 28). Нога слугує для заривання в ґрунт і повільного повзання. У молюсків, що ведуть сидячий спосіб життя, нога значно редукується і може зовсім зникнути, як в устриць. У багатьох двостулкових на нижній поверхні ноги в особливій заглибині є так звана бісусна залоза, яка виділяє тягучу органічну речовину, що твердне у воді, перетворюючись на міцні нитки — бісус, за допомогою якого молюск прикріплюється до субстрату. Така бісусна залоза є, наприклад, у мідії (Mytilus), дрейсени (Dreissena) тощо.
Покриви двостулкових утворені шкірним епітелієм із великою кількістю слизових клітин та сполучною тканиною з окремими м’язовими волокнами й лакунами, заповненими гемолімфою. Епітелій у більшій частині війчастий (епітелій ноги, внутрішньої стінки мантії, зябер, ротових лопатей).
Мускулатура двостулкових поділяється на дві групи м’язів: одна — пов’язана з ногою, друга — з мантією та черепашкою (рис. 30). Нога має здебільшого дві (у Nuculidae — чотири) пари ретракторів, які прикріплюються до черепашки й при скороченні втягують ногу (див. рис. 25, в). Крім того, у нозі є безліч м’язових волокон, які розходяться у різних напрямах і утворюють майже справжній шкірно-м’язовий мішок, що спирається на кров’яний синус усередині ноги. Перед висуванням ноги в її синус нагнітається гемолімфа, яка при втягуванні ноги із синуса перекачується в центральний венозний синус, і об’єм ноги різко скорочується. Про м’язи мантії та черепашки вже говорилося.
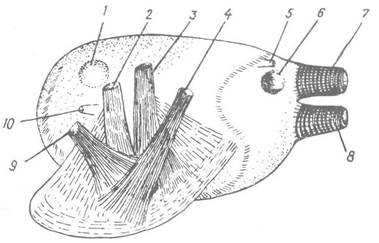
Рис. 30. Мускулатура Bivalvia:
1 — передній м’яз-замикач; 2,4 — ретрактори ноги; 3 — леватор (підіймач) ноги; 5 — анус; 6 — задній м’яз- замикач; 7, 8 — кільцеві та поздовжні м’язи сифонів; 9 — протрактор ноги; 10 — рот
Вторинна порожнина тіла (целом) у двостулкових складається з дуже невеличкого перикардія та порожнин гонад. Решта проміжків між внутрішніми органами заповнені сполучною тканиною —паренхімою, багатою на лакуни.
Травна система починається на передньому кінці тіла ротовим отвором, оточеним з боків двома парами ротових лопатей (ротові щупальця). З редукцією голови в двостулкових пов’язана відсутність у травному тракті глотки, радули, щелеп і слинних залоз.
Рот веде в короткий стравохід (ектодермальна передня кишка), вистелений миготливим епітелієм. Він відкривається в мішкоподібний шлунок, який належить до ентодермальної середньої кишки. Шлунок оточує парна печінкова залоза, яка складається з безлічі дрібних часток, що відкриваються кількома (їх може бути дві, три або багато) протоками в шлунок (рис. 31, а).
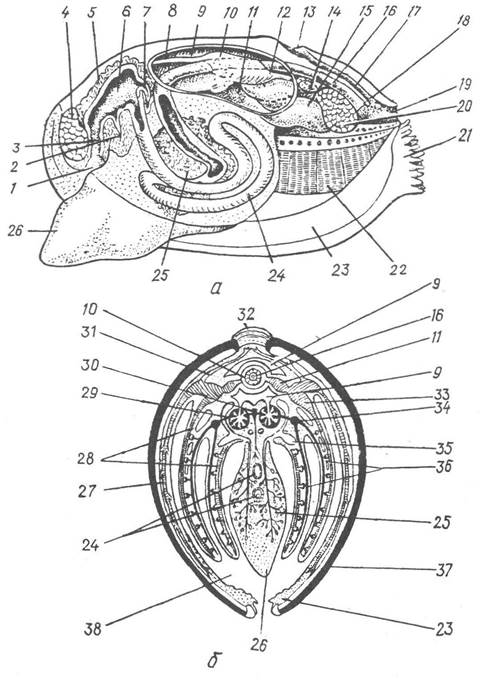
Рис. 31. Схема внутрішньої будови беззубки:
а — поздовжній розріз через тіло, вигляд зліва; б — поперечний розріз та схема кровоносної системи; 1 — рот; 2 — мішок кришталевого стовпчика; 3 — стравохід; 4 — передній м’яз-замикач; 5 — печінка; 6 — шлунок; 7 — протока печінки; 8 — передня аорта; 9 — перикардій; 10 — шлуночок серця; 11 — передсердя; 12 — задня аорта; 13 — спинний мантійний отвір; 14 — задній ретрактор ноги; 15 — нирка; 16 — задня кишка; 17 — спинний мантійний канал; 18 — анальний отвір; 19 — вивідний сифон; 20 — задній м’яз-замикач; 21 — ввідний сифон; 22 — напівзябра; 23 — мантійний листок; 24 — середня кишка; 25 — гонада; 26 — нога; 27 — мантійна вена; 28 — виносні зяброві судини; 29 — венозна система нирок; 30 — нутрощева та ножна вени; 31 — судини від кеберового органа; 32 — лігамент; 33 — зяброва вена; 34 — зяброва артерія; 35 — верхня камера мантійної порожнини; 36 — приносні зяброві судини; 37 — стулка черепашки; 38 — нижня камера мантійної порожнини
У задню частину шлунка відкривається сліпе мішкоподібне вип’ячування, у порожнині якого утворюється склоподібно-прозора драглиста паличка — кришталевий стовпчик (рис. 32). Це неклітинний гіаліновий стрижень, до складу якого входять мукопротеїни та ферменти, що розщеплюють вуглеводи (амілаза, глікогеназа тощо). Вільний кінець кришталевого стовпчика впирається в так званий гастричний (шлунковий) щит — невеличку прозору пластинку ущільненого секрету, яка міститься на передній стінці шлунка.
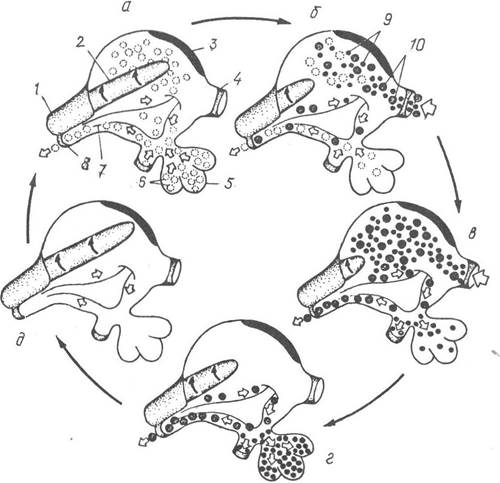
Рис. 32. Цикл травлення в шлунку та травній залозі (печінці) двостулкових молюсків:
а — шлунок перед прийняттям їжі; б — надходження їжі до шлунка та початок травлення і сортування; в — сортування харчових частинок у шлунку; г — внутрішньоклітинне травлення дрібних частинок у печінці та видалення великих частинок у кишку; д — шлунок порожній, підготовка до нового циклу; 1 — мішок кришталевого стовпчика; 2 — кришталевий стовпчик; 3 — гастричний щит; 4 — стравохід; 5 — печінка; 6 — фрагменти травних клітин з ферментами; 7 — великий тифлозоль;
8 — початок середньої кишки; 9, 10 — дрібні та великі харчові частинки
Більша частина внутрішньої поверхні шлунка має борозенчасті зони, вкриті війками і розділені вузькими провідними жолобками. Це сортувальні зони, де відбувається сортування харчових частинок.
Від шлунка відходить середня кишка, стінки якої утворюють дві довгі внутрішні складки — тифлозолі; часто тифлозолі вдаються в порожнину шлунка. Стінки середньої кишки також вистелені війчастим епітелієм. Середня кишка спускається від шлунка до основи ноги, утворює в ній кілька петель і прямує до спинної сторони тулуба, де переходить у задню кишку. Задня кишка звичайно пронизує шлуночок серця і закінчується порошицею на дні вивідного сифона (див. рис. 31, а).
Живляться двостулкові рослинним детритом та дрібними організмами, що завжди є в мулі, або придонними планктонними організмами, які молюски пасивно відфільтровують із води, що проходить через їх мантійну порожнину. Усе це зліплюється слизом у грудочки і прямує до харчових жолобків на нижніх краях зябер, а по них — до ротових лопатей. Ротові лопаті також вкриті війками і мають борозенки, по яких харчові грудочки просуваються до ротового отвору. Неїстівні частинки, так звані псевдофекалії, викидаються назовні.
Лише деякі морські форми є хижаками (переважно з надряду Septibranchia). Для молюсків-деревоточців їжею є, крім того, деревина. Отже, більшість двостулкових живиться дисперсними частинками, які збираються поверхнею зябер і ротових лопатей.
їжа надходить до шлунка із стравоходу у вигляді стрічкоподібного слизового шнура з харчовими частинками. Завдяки биттю війок мішка, в якому міститься кришталевий стовпчик, він обертається навколо своєї осі й працює як коловерт, витягуючи слизовий шнур із стравоходу. Водночас при обертанні стовпчика, який треться об поверхню гастричного щита, утворюється суспензія із частинок їжі та рідини з ферментами, які вивільнюються при розчиненні кінчика кришталевого стовпчика в слабокислому середовищі шлунка. Далі ця суспензія потрапляє до сортувальних зон, де частинки діляться за розміром та вагою. Дрібні, легкі частинки переносяться до печінкових проток і втягуються в них. У печінці вони фагоцитуються і перетравлюються печінковими клітинами. Більші частинки їжі із шлунка прямують в середню кишку і згодом викидаються з екскрементами. Фагоцитоз відбувається також і в кишечнику. В епітелії кишечника є амебощні клітини, здатні виповзати в просвіт середньої кишки, поглинати харчові частинки і потім повертатися до стінки кишки. Вуглеводи перетравлюються позаклітинно, у порожнині шлунка за допомогою ферментів кришталевого стовпчика, а білки й жири — переважно внутрішньоклітинно в печінці й середній кишці.
У хижих Septibranchia в шлунку немає кришталевого стовпчика та сортувальних зон; їх шлунок має добре розвинені м’язові стінки, а його епітелій вистелений хітиноїдною кутикулою. їжа (переважно дрібні ракоподібні) в такому шлунку розчавлюється й перетирається, а перетравлюється в печінці.
Видільна система Bivalvia складається з пари нирок, або боянусових органів. Вони лежать під перикардієм і мають вигляд мішкоподібних трубок, кожна з яких одним кінцем відкривається в перикардій, а другим — у мантійну порожнину біля основи ноги (див. рис. 31, я). У різних двостулкових форма нирок буває різною: найчастіше нирки складені вздовж, тому набувають V-подібної форми, у деяких вони S-подібно вигнуті. Отвір нирки в перикардій має клапан — невеличку складку, що перешкоджає зворотній течії рідини. Далі йде лійка — короткий тонкостінний каналець, вистелений миготливим епітелієм. За нею міститься залозистий відділ, внутрішня поверхня якого вистелена залозистим епітелієм і утворює численні складки, які вдаються в порожнину нирки. У них є густа мережа венозних лакун, проходячи якими гемолімфа залишає значну кількість продуктів дисиміляції. Залозистий відділ переходить у тонкостінний каналець, через який екскрети виводяться в мантійну порожнину.
Видільну функцію виконує також так званий кеберів орган, що є розростанням передньої стінки перикардія (див. рис. 25, в). Залозисті клітини кеберового органа вилучають із гемолімфи, яка омиває перикардій, продукти метаболізму і виділяють їх у порожнину останнього, звідки через нирки виводять їх назовні.
Кровоносна система в двостулкових незамкнена, складається із серця, кровоносних судин та системи лакун і синусів. Серце міститься на спинній стороні тіла і має здебільшого один шлуночок та два бічних передсердя (див. рис. 31). Воно оточене навколосерцевою сумкою —перикардієм, який утворений целомічним (перитонеальним) епітелієм. Кожне передсердя сполучається із шлуночком отвором, перекритим складчастим клапаном. Завдяки цьому при скороченні передсердь гемолімфа прямує лише від передсердь до шлуночка. Крізь шлуночок серця в більшості двостулкових проходить задня кишка, але порожнини їх не сполучаються. Від шлуночка до переднього і заднього кінців тіла відходять передня та задня аорти, які поділяються на артерії, що галузяться й врешті-решт відкриваються в лакуни.
Циркуляція гемолімфи забезпечується роботою серця й м’язів тіла. Насичена киснем гемолімфа з передсердь надходить у шлуночок серця, звідти виштовхується в судини, далі — в лакуни, де віддає кисень тканинам і насичується вуглекислим газом. З тканин венозна гемолімфа збирається в центральний венозний синус під перикардієм. Потім потрапляє до видільної системи, де звільнюється від продуктів обміну, а звідти переганяється в зябра. Зябровий апарат має складну систему судин і лакун, де гемолімфа збагачується киснем. Із зябер гемолімфа надходить у зяброві вени, які відкриваються в передсердя (див. рис. 31, б). Крім того, значна частина гемолімфи насичується киснем у мантії та ротових лопатях, які відіграють неабияку роль у газообміні; з цих органів по мантійних венах гемолімфа надходить також у зяброві вени.
Як уже згадувалось, у більшості двостулкових крізь шлуночок серця проходить задня кишка, проте в деяких форм серце лежить над кишкою або під нею. Це пояснюється тим, що під час ембріонального розвитку парний мезодермальний зачаток, з якого утворюються перикардій, серце й нирки, спочатку міститься обабіч кишки, а потім обидві його половини з’єднуються над і під кишкою, охоплюючи її у вигляді муфти, але в деяких форм таке злиття відбувається тільки над або під кишкою.
Гемолімфа двостулкових містить багато клітин, серед яких найбільше поширені різні типи амебоцитів; у деяких видів є й еритроцити. Гемолімфа може бути безбарвною (наприклад, у Anodonta), або червоною через наявність червоного пігменту гемоглобіну, який міститься в плазмі чи еритроцитах. Гемоглобін виконує функцію запасання кисню, який віддає в періоди, коли в оточуючому середовищі його мало (наприклад, при зариванні молюска в мул).
Гемолімфа двостулкових виконує різноманітні функції. Вона забезпечує сталість іонного складу та осмотичного тиску внутрішнього середовища організму; постачає тканинам кисень і поживні речовини; видаляє продукти обміну та вуглекислий газ. Крім того, амебоцити, зосереджені в лакунах нирок, накопичують у цитоплазмі кристалики екскретів і виносять їх у порожнину нирок. Захисна функція гемолімфи зумовлена тим, що амебоцитам притаманні фагоцитарні властивості. Так, при запальних явищах ці клітини енергійно фагоцитують мертві клітини, бактерій, а при паразитуванні личинок деяких трематод навколо них утворюється капсула з цих клітин. Нарешті, при пораненні клітинні елементи гемолімфи аглютинують, що припиняє кровотечу й призводить до загоєння рани. Гемолімфа двостулкових, як і інших молюсків, виконує ще функцію гідроскелета при рухах ноги. Висування ноги відбувається при розслабленні її м’язів і надходженні до лакун гемолімфи, а втягування — при скороченні мускулатури і відтіканні гемолімфи в черевний венозний синус.
Органами дихання двостулкових є пара зябер, які розташовані в мантійній порожнині обабіч ноги. У примітивних форм (надряд Protobranehia) —це типові двопірчасті ктенідії, які складаються з осі та двох рядів трикутних зябрових пелюсток, що звисають у порожнину мантії (рис. 33, а). У більшості двостулкових (надряд Autobranchia) у зв’язку з переходом до живлення шляхом фільтрації води, зябра перетворилися на органи фільтрації і набули вигляду великих пластин, що займають більшу частішу мантійної порожнини. У більшості Autobranchia, у тому числі й у прісноводних перлівниць, кожна із зябер складається з двох пластинчастих півзябер: зовнішньої, оберненої до мантії, і внутрішньої, оберненої до ноги. Півзябра складаються з численних тонких трубочок — зябрових ниток (філаментів), які утворюють низхідне (внутрішнє) та висхідне (зовнішнє) коліна. Філаменти з’єднуються між собою короткими поперечними сполучнотканинними перемичками, а висхідні та низхідні коліна кожного філамента сполучаються численними септами (рис. 33, в). Отже, кожна півзябра є двошаровою пластинкою, продірявленою дрібними порами, які ведуть до системи щілин у внутрішньозябровій порожнині. Проте в деяких молюсків (наприклад, у мідій) сусідні зяброві філаменти з’єднуються один із одним лише війками, тому зяброві пластинки легко розтріпуються на окремі нитки (рис. 33, б). Зяброві пластинки зверху і зсередини вкриті війчастим епітелієм, між епітеліальними клітинами містяться залозисті клітини, які виділяють слиз. Зябра зміцнюються сітчастим опорним «скелетом» — у кожному філаменті під епітелієм є потовщення сполучної тканини, тобто тонкі хітиноїдні палички.
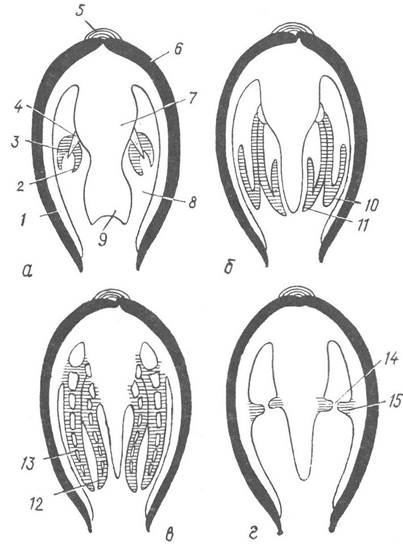
Рис. 33. Будова зябер представників трьох надрядів двостулкових (схематичні поперечні зрізи):
а — Protobranchia; б—в — Autobranchia; г — Septibranchia; 1 — мантія; 2 — внутрішній листок ктенідія; 3 — зовнішній листок ктенідія; 4 — вісь ктенідія; 5 —лігамент; 6 — черепашка; 7 — тулуб; 8 — мантійна порожнина; 9 — нога; 10 — зовнішня зяброва нитка; 11 — внутрішня зяброва нитка; 12 — внутрішня напівзябра; 13 — зовнішня напівзябра; 14 — пора в зябровій септі; 15— зяброва
септа
Над кожною півзяброю від переднього кінця тіла до заднього тягнуться надзяброві канали, пов’язані з вивідним сифоном.
Узгоджена дія війок миготливого епітелію зябер, ротових лопатей, мантії й покривів тіла створює різницю гідростатичних тисків у надзябровій та підзябровій порожнинах, а також в останніх та зовнішньому середовищі. Внаслідок цього вода надходить через ввідний сифон або через широкий педальний отвір у мантійну порожнину, омиває зябра, через пори потрапляє до каналів, розташованих всередині зябер; із них, віддавши кисень і збагатившись вуглекислим газом, вода потрапляє в надзяброві канали і звідти викидається назовні через вивідний сифон (рис. 34).
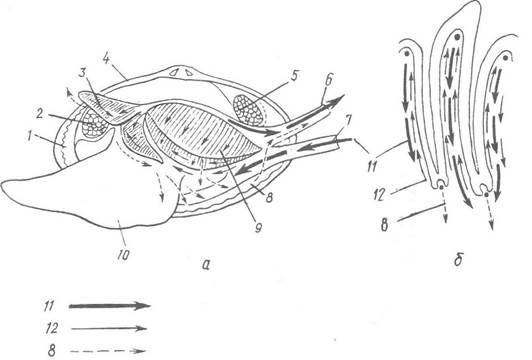
Рис. 34. Фільтрація води справжніми пластинчастозябровими (надряд Autobranchia) для дихання та живлення:
а — загальна схема; б — струми води на зябрах мідії; 1 — мантія; 2 — передній м’яз-замикач; 3 — ротові лопаті; 4 — черепашка; 5 — задній м’яз-замикач; б — вивідний сифон; 7 — ввідний сифон; 8 — шляхи псевдофекалій (неїстівних частинок); 9 — зябра; 10 — нога; 11 — напрям основних токів води, які надходять у мантійну порожнину та виходять з неї; 12 — фільтраційні та сортувальні струми
на зябрах та ротових лопатях
У деяких двостулкових (підклас Septibranchia) ктенідії атрофуються, і в мантійній порожнині утворюється мускульна перетинка, яка поділяє її на дві ділянки —нижню та верхню, в якій і здійснюється газообмін (див. рис. 33, г).
Нервова система двостулкових розкидано-вузлового типу, вона складається з трьох пар нервових гангліїв: головних (цереброппевральних), ножних (педальних) і тулубних (вісцеропарієтальних), з’єднаних між собою комісарами й конективами. Цереброплевральні ганглії лежать на передньому кінці тіла, здебільшого по боках від ротового отвору, обидва ганглії з’єднуються над стравоходом церебральною комісурою (рис. 35). Від них відходять нерви до ротових лопатей, передніх м’язів-замикачів та передньої частини мантії. У нозі залягає пара педальних гангліїв, які з’єднуються з цереброплевральними за допомогою двох довгих конектив. Ганглії зближені між собою і зв’язані короткою комісурою; педальні ганглії іннервують ногу. Під заднім м’язом-замикачем залягає пара тісно зближених вісцеропарієтальних гангліїв. Вони іннервують задню частину мантії, задні м’язи-замикачі, зябра, осфрадії та нутрощі. Довгими конективами вони з’єднані з цереброплевральними гангліями, а в деяких видів — і з педальними.
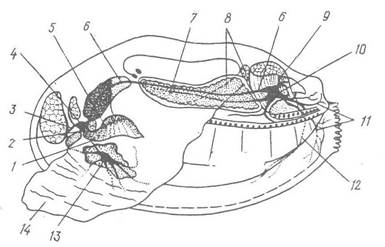
Рис. 35. Нервова система Anodonta (вигляд з лівої сторони):
1 — цереброплевропедальна конектива; 2 — церебральна комісура; 3 — нерв переднього м’яза замикача; 4 — цереброплевральний ганглій; 5 — нерв ротової лопаті; 6 — цереброплевровісцеральна конектива; 7 — нирка; 8 — задній ретрактор ноги; 9 — нерв заднього м’яза-замикача; 10 — вісцеропарієтальний ганглій; 11 — нерви сифонів; 12 — мантійний нерв; 13 — педальний ганглій;
14 — нога
Відсутність голови, спрощення ротового апарату і малорухливий спосіб життя двостулкових зумовили слабкий розвиток у них органів чуття. Органів чуття, пов’язаних в інших молюсків з головою (очі, головні щупальця), у них немає.
Проте досить часто буває, що органи зору вторинно виникають або по всьому краю мантії (гребінець — Pecten), або по краю сифонів (серцевидка — Cardium), і навіть на зябрах (арка — Arca). Такі вторинні очі мають різну будову — від очних ямок до таких відносно складно побудованих очей з кришталиком та сітківкою, як мантійні очі гребінців або тридакн ( рис. 36).
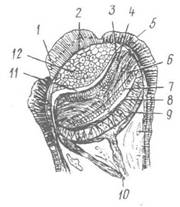
Рис. 36. Розріз мантійного ока Pecten islandicus:
1 — рогівка; 2 — сполучнотканинний шар; 3 — очна септа; 4 — дистальний шар сітківки; 5 — зовнішній епітелій ока; 6 — зорові клітини; 7 — шар світлочутливих паличок; 8 — тапетум; 9 — пігментний шар; 10 — оптичний нерв; 11 — лакунарний простір; 12 — кришталик
Органи дотику представлені чутливими клітинами, розкиданими по всій поверхні тіла — найбільше їх на ротових лопатях, нозі, краях сифонів та мантії.
Органи хімічного чуття — осфрадії та пов’язані з ними чутливі валки — є утвореннями, вкритими чутливими нервовими клітинами. їх функція —контроль якості води, яка надходить до мантійної порожнини. Вони є в різних місцях — на нозі, складках мантії, біля зябер, сифонів.
Органи рівноваги — статоцисти— у більшості двостулкових містяться в нозі, іноді — на спинній стороні тіла. Здебільшого їх два. Це пухирці, стінки яких складаються з чутливого епітелію, а всередині, у рідині, що їх заповнює, міститься один масивний статоліт або численні дрібні піщинки — статоконії.
Переважна більшість двостулкових роздільностатеві, лише деякі групи — гермафродити, наприклад прісноводні кулькові (родина Sphaeiiidae). Деяким двостулковим властива зміна статі протягом життя (морські перлові скойки). Гонади парні й містяться в основі ноги. Вони мають вигляд гроноподібних, розгалужених органів; порожнина їх є целомічним утворенням. При сильному розвитку гонади вирости можуть заходити в тканини ноги (наприклад, у беззубки, див. рис. 31), або мантії (у мідії). У більшості двостулкових від гонад відходять коротенькі статеві протоки, що відкриваються в мантійну порожнину статевими отворами, які лежать поблизу від отворів нирок. Лише в деяких форм (надряд Protobranchia, а також роди Pecten, Ostrea) гонади не мають власних проток і відкриваються у нирки. Копулятивних органів немає, запліднення яєць переважно зовнішнє.
Більшість двостулкових відкладає яйця у воду, де й здійснюється запліднення. Проте багато прісноводних форм (родина Unionidae) відкладають яйця в зябра, де відбувається запліднення й розвиток до виходу личинки. У деяких видів проявляється турбота про нащадків. Самиці Cardium elegantulum відкладають яйця в дві виводкові сумки, утворені на стінках мантії (рис. 37, а), тут проходить розвиток зародка, без планктонної личинки. У дельфінозубої горіховидки (Nucula delphinodonta) яйця розвиваються в зовнішній шкірястій сумці, яка прикріплена до черепашки і сполучається з мантійною порожниною (рис. 37, б). Деякі двостулкові є живородними (наприклад, прісноводні Sphaeriidae та Pisidiidae).
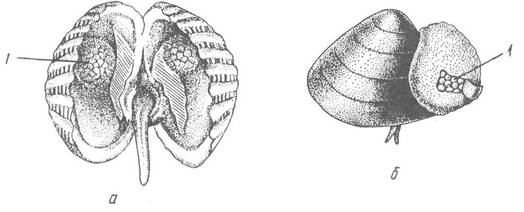
Рис. 37. Виводкові сумки двостулкових:
а — розкрита черепашка Cardium elegantulum з двома виводковими сумками, заповненими зародками; б — Nucula delphinodonta із зовнішньою виводковою сумкою; в — зародки
У результаті спірального дробіння яйцеклітини утворюється личинка трохофорного типу (рис. 38, а). У багатьох видів трохофора має зачаток ноги й первинну черепашку, яка спочатку закладається у вигляді однієї пластинки на спинній стороні. Згодом трохофора перетворюється на велігера — личинку, яка має вкритий війками диск — парус (велум), двостулкову черепашку, зачатки внутрішніх органів (рис.38, б). За допомогою цих личинок, що ведуть планктонний спосіб життя і переносяться течіями, малорухливі молюски розселюються. У прісноводних уніонід з яєць, відкладених у зяброві порожнини самиць, виходять своєрідні личинки — глохідії (рис. 39), які деякий час паразитують на рибі (див. с. 56).
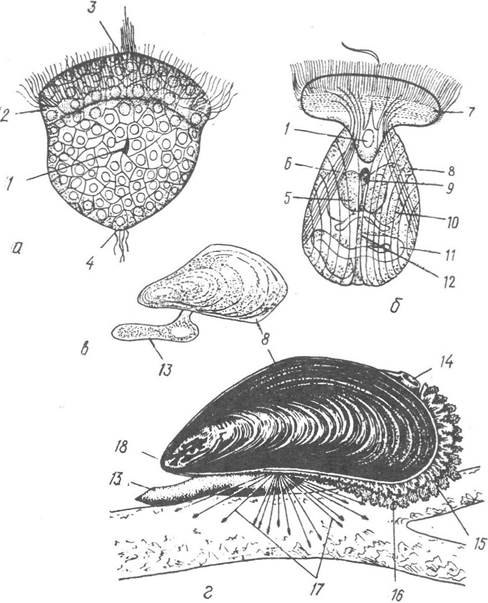
Рис. 38. Метаморфоз мідії:
а — трохофора; б — велігер; в — повзаюча личинка; г — доросла мідія; 1 — рот; 2 — прототрох; З — тім’яна пластинка; 4 —телотрох; 5 — педальний ганглій; 6 — ретрактор прототроха; 7 — парус; 8 — черепашка; 9 — вхід до мантійної порожнини; 10 — печінка; 11 — кишечник; 12 — анальний отвір; 13 — нога; 14 — вивідний сифон; 15 — ввідний сифон; 16 — фестончастий край мантії; 17 — нитки бісусу; 18 — передній кінець черепашки
Двостулкові молюски відіграють важливу роль у морських та прісноводних біоценозах. Як дорослі м’якуни, так і їхні личинки є важливими ланками в ланцюгах живлення. Дорослих двостулкових споживають морські зірки, хижі черевоногі та головоногі молюски, риби, птахи, ссавці. Личинками двостулкових живляться всі планктонні фільтруючі організми. Двостулкові є найефективнішими фільтраторами, які очищають воду.
Практичне значення двостулкових зумовлене тим, що багато їх видів людина здавна споживає як висококалорійну та смачну їжу (устриці, мідії, морські гребінці тощо); крім того, черепашки деяких молюсків використовують для виготовлення прикрас і гудзиків, для інкрустаційних робіт. Перлові скойки дають цінні перла. Проте деякі види молюсків приносять шкоду людині, наприклад дрейсена, якою обростають днища суден і тим уповільнюється їх хід; вона оселяється в турбінних решітках електростанцій, у трубах водопроводів тощо, перешкоджаючи їх роботі.
В основі системи класу Двостулкові лежить будова черепашки, особливо її замка, та зябер. Останнім часом до діагностичних ознак відносять і деякі риси внутрішньої будови, зокрема будову шлунка. Клас Bivalvia поділяється на три надряди — Первиннозяброві (Protobranchia), Пластинчастозяброві (Autobranchia) та Перетинчастозяброві (Septibranchia):
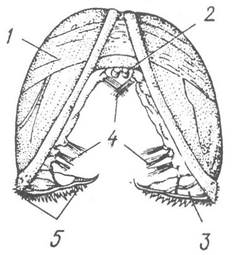
Рис. 39. Глохідій Anodonta:
1 — черепашка; 2 — личинкова бісусна залоза; 3 — крайовий зубець; 4 — пучки чутливих щетинок; 5 — зубчики на крайовому зубці