Зоология позвоночных - В. М. Константинов 2011
Хордовые
Челюстноротые
Класс Костные рыбы - Подкласс Лопастепёрые рыбы - Надотряд Кистепёрые рыбы
Кистепёрые — древняя и почти вымершая группа рыб. Сравнительно широкое распространение кистепёрые имели в девоне и карбоне. Их ископаемые остатки в соответствующих слоях обнаружены в пресных и морских водах всей планеты, от Шпицбергена и Новой Земли до Южной Африки, Австралии и Антарктиды. В мезозое число видов и широта распространения их сократились. Первый экземпляр этих удивительных рыб был добыт в 1938 г. в Индийском океане, у южных берегов Африки, близ устья реки Халумна, на глубине около 70 м. Это была крупная рыба длиной 150 см и массой 57 кг. Она была названа латимерией (Latimeria chalumnae) по имени госпожи Латимер, смотрительницы местного краеведческого музея, обнаружившей удивительную рыбу в улове траулера и сохранившую ее для науки. В последующем в этом районе неоднократно ловили кистепёрых рыб. Местное население ловит их систематически и употребляет в пищу (рис. 46).

Рис. 46. Латимерия
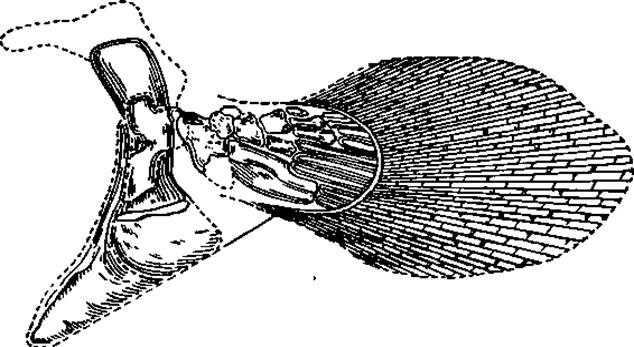
Рис. 47. Скелет плавника кистепёрой рыбы (Sauripterus)
Все современные кистепёрые — латимерии, или целоканты, обнаружены только в районе Коморских островов, где они держатся на глубине 400 — 1 000 м при температуре воды 10... 14°С. Длина тела половозрелых особей 125 — 180 см, масса 25 — 80 кг. Позвонки зачаточны, пожизненно хорошо развита хорда. Первичный череп в значительной мере хрящевой. Целоканты — хищники: их рот вооружен многочисленными острыми зубами.
В полости тела обнаружено окруженное жиром дегенерировавшее легкое. Внутренних ноздрей у целокантов нет, и они в отличие от мезозойских кистепёрых не способны к дыханию атмосферным кислородом.
Тело кистепёрых покрыто чешуями, представляющими собой толстые костные пластинки округлой или ромбической формы, одетые сверху слоем видоизмененного дентина и тонким слоем эмали. Парные плавники имеют своеобразное устройство. В основании их расположена широкая мясистая (мышечная) лопасть, внутри которой находится скелет основной части самого плавника (рис. 47). К настоящему времени поймано около 100 латимерий.
Кистепёрые, несомненно, близки к двоякодышащим и возникли, видимо, от одного с ними корня. Они обитали первоначально в пресных водоемах, где в засаде поджидали добычу. Периодически, вероятно, они испытывали недостаток кислорода: в связи с этим развилось двойное дыхание. Засоренность водоемов отмершей растительностью явилась, видимо, предпосылкой для развития своеобразных парных конечностей, которые имели мускулатуру на самой Конечности и могли использоваться не только для гребли, но и для опоры о твердый субстрат: дно, стебли, стволы деревьев.
Развитие древних кистепёрых рыб пошло по крайней мере двумя самостоятельными путями. Один из них — возникновение целокантов (отряд Целокантообразные — Caelocantiformes) — привел этих рыб в океан. Представители этой линии развития дожили до настоящего времени и предстали в облике латимерии. Другая группа кистепёрых — Рипидистиеобразные (Rhipidistiiformes) — в одной из своих эволюционных линий развития (остеолепиформные рипидистии) сочетала в своей организации черты приспособления к дыханию в наземной среде (легкие, внутренние ноздри — хоаны) и к движению по твердому субстрату (унисериальный тип парных плавников, близкий к пятипалой конечности). Вероятно, их потомками стали наземные позвоночные.
ЭКОЛОГИЯ РЫБ
Некоторые условия жизни рыб в водной среде
Жизненная арена рыб исключительно велика. При общей поверхности земного шара, равной примерно 510 млн км2, 71 %, или около 361 млн км2 всей площади, занято поверхностью океанов и морей. Около 2,5 млн км2, или 0,5 % площади земного шара, занято внутренними водоемами. Обширность жизненного пространства рыб определяется, кроме того, и большим простиранием его по вертикали. Максимальная известная глубина океана равна примерно 11 тыс. м. Океаны с глубиной более 3 тыс. м занимают примерно 51 —58 % от всей площади морских вод. Рыбы живут в областях, расположенных от экватора до приполюсовых пространств; они есть в горных водоемах на высоте более 6 тыс. м над уровнем моря и в океанах на глубине более 10 тыс. м. Все это создает большое разнообразие условий существования.
Вода как жизненная среда обладает рядом специфических особенностей, создающих своеобразные условия для обитания в ней животных, и способствует их широчайшему видовому и экологическому многообразию.
Рассмотрим некоторые черты водной среды применительно к населяющим ее рыбам.
Подвижность водной среды. Подвижность водной среды связана с постоянными течениями в реках и морях, местными течениями в мелких замкнутых водоемах, вертикальными перемещениями слоев воды, вызываемыми различным их прогреванием.
Подвижность воды обусловливает в значительной мере пассивные перемещения рыб. К примеру, личинки норвежской сельди, которые выводятся у берегов Западной Скандинавии, увлекаются одной из ветвей течения Гольфстрим на северо-восток и за 3 месяца относятся вдоль побережья на 1 000 км (рис. 48). Мальки многих лососевых рыб выводятся в верховьях притоков крупных рек, а основную часть жизни проводят в морях. Переход из рек в моря совершается также в значительной мере пассивно благодаря течениям рек. Наконец, подвижность воды обусловливает пассивные передвижения кормовых объектов — планктона, что, в свою очередь, влечет за собой и перемещение рыб.
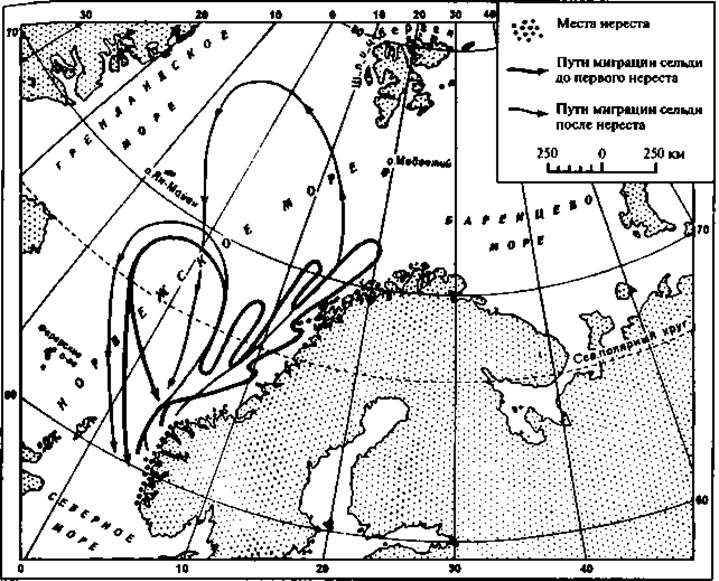
Рис. 48. Схема миграции норвежской сельди
Колебания температуры. Колебания температуры в водной среде значительно меньшие, чем в воздушно-наземной. В подавляющем большинстве случаев верхний предел температуры, при которой встречаются рыбы, лежит ниже 30 —40°С. Особенно характерен нижний предел температуры воды, который даже в сильно соленых частях океанов не опускается ниже -2°С. Следовательно, реальная амплитуда температур среды обитания рыб 35—45°С. Однако и эти сравнительно ограниченные колебания температуры имеют в жизни Рыб большое значение.
Рыбы принадлежат к пойкилотермным животным. Температура их телa находится в прямой зависимости от температуры окружающей среды и меняется вслед за ее изменениями. Температура воздействует на организм рыб как прямо, так и косвенно — благодаря способности воды растворять газы и меняя газообмен.
Наличие кислорода в воде. Известно, что способность воды растворять газы, в частности кислород, обратно пропорциональна ее температуре и солености. Это видно из данных табл. 4. Вместе с тем по мере повышения температуры воды потребность рыб в кислороде увеличивается.
Выявлена минимальная концентрация кислорода, ниже которой рыба погибает. Для карпа при температуре 1°С она будет равна 0,8 мг/л, при 30°С — 1,3 мг/л, а при 40°С — около 2,0 мг/л.
Неодинакова и потребность разных видов рыб в кислороде. По этому признаку они могут быть разбиты на четыре группы: 1) требующие очень много кислорода — 7—11 см3/л: кумжа (Salmo trutta), гольян (Phoxinus phoxinus), арктический голец (Salvelinus alpinus), 2) требующие много кислорода — 5 — 7 см3/л: хариус (Thymallus thymallus), голавль (Leuciscus cephalus), пескарь (Gobio gobio); 3) потребляющие сравнительно небольшое количество кислорода — около 4 см3/л: плотва (Rutilus rutilus), окунь (Perca fluviatilis), ерш (Acerina cernuus); 4) выдерживающие очень слабое насыщение воды кислородом и живущие даже при 0,5 см3/л: сазан, линь, карась. Отсюда и постоянная потребность в обновлении воды для одних видов, и способность других жить в относительно спокойных, слаботекущих или стоячих водоемах.
Огромное значение в жизни рыб, как положительное, так и отрицательное, имеет зимнее сезонное образование льда в водоемах.
Ледяной покров изолирует нижележащие слои воды от низких температур воздуха и тем самым предотвращает промерзание водоема до дна. (Лишь иногда мелководные водоемы промерзают до дна.) Это делает возможным распространение рыб в областях с очень низкими зимними температурами воздуха. Таково положительное значение ледяного покрова. Однако покров льда играет в жизни рыб и отрицательную роль. Он затемняет водоемы, а это замедляет и даже приостанавливает жизненные процессы у многих водных организмов, прямо или косденно имеющих для рыб пищевое значение. В первую очередь это касается зеленых водорослей и высших растений, которыми кормятся сами рыбы и те беспозвоночные животные, которые служат рыбам пищей.
Таблица 4
Количество кислорода в воде (см3/л) при разной температуре и солености
Температура воды, °С |
Соленость воды, мг/л |
||
0 % |
20 % |
35 % |
|
0 |
10,3 |
9,0 |
8,0 |
10 |
8,0 |
7,1 |
6,4 |
20 |
6,6 |
5,9 |
5,4 |
30 |
5,6 |
5,0 |
4,5 |
Ледяной покров резко снижает возможность пополнения воды кислородом из воздуха. Зимой во многих водоемах в результате процессов гниения полностью расходуется кислород, растворенный в воде. Возникает явление, известное под названием «замора» рыбы. В России оно имеет широкое распространение и происходит в бассейнах, водосборная площадь которых связана в значительной мере с торфяными болотами. Большие заморы существуют, например, в бассейне реки Обь. Болотные воды, питающие здесь реки, богаты гуминовыми кислотами и оксидами железа. Последние, окисляясь, отбирают из воды растворенный в ней кислород. Возмещение же его из воздуха невозможно из-за сплошного ледяного покрова, надолго нарушающего жизнь водоемов.
Заморы бывают и в реках европейской части России. С ними ведут успешную борьбу путем создания прорубей, усиления проточности прудов и озер. В прудовых хозяйствах с высокой технической оснащенностью употребляют компрессоры, нагнетающие воду, обогащенную кислородом.
Звукопроводимость. Еще одно свойство воды — звукопроводимость. Это явление широко используется рыбами для получения всесторонней информации о пище, хищниках, об аномальных явлениях. Среди рыб хорошо развита звуковая сигнализация. Она обеспечивает обмен информацией как между особями одного вида, так и между особями разных видов. Издаваемые рыбами звуки имеют безусловно и эхолокационное значение (см. подраздел об органах чувств).
Жизненный цикл рыб. Миграции
Жизненный цикл рыб складывается из ряда последовательно сменяющих друг друга этапов: созревания, размножения, нагула, зимовки. Во время каждого из этапов жизненного цикла рыбы нуждаются в специфических условиях среды, которые они находят в различных, зачастую далеко отстоящих друг от друга местах водоема, а иногда и в разных водоемах.
Так, условия, необходимые для нереста, отличны от условий, обеспечивающих наилучший нагул рыб. Своеобразные требования предъявляют рыбы к условиям зимовки и т. д. Это приводит к тому, что в поисках мест, подходящих для каждого жизненного отправления, рыбы совершают более или менее значительные перемещения. У видов, населяющих мелкие замкнутые водоемы (пруды, озера) или Реки, перемещения имеют ничтожные масштабы, хотя и в этом случае они все же выражены вполне отчетливо. Выделяют нерестовые, кормовые, зимовальные миграции.
Степень развитости нерестовых миграций неодинакова у разных видов. Некоторые рыбы, и их большинство, ежегодно (или через некоторые интервалы) нерестятся, повторяя одни и те же передвижения. Другие в течение жизненного цикла только один раз проходят стадию созревания половых продуктов, один раз предпринимают нерестовую миграцию и только однажды в жизни размножаются. Таковы некоторые виды лососевых (кета, горбуша), речные угри.
Особенно сильно развиты миграции у морских, тем более у проходных рыб. Многие чисто морские рыбы совершают в связи с нерестом длительные перемещения, переходя из открытого моря к берегам или, наоборот, от прибрежий в глубину моря. Таковы морские сельди, треска, пикша и др. Длина миграционного пути зависит от того, как далеко от мест нагула расположены места, подходящие для нереста.
Чрезвычайно сложны и разнообразны нерестовые миграции у проходных рыб: они связаны с переходом из морей в реки — анадромные миграции (чаще) или, наоборот, из рек в моря (реже) — катадромные миграции.
Переход для размножения из морей в реки свойствен многим лососевым, осетровым, некоторым сельдям и карповым. Значительно меньше видов, кормящихся в реках и для нереста идущих в моря. Таковы, например, угри.
Протяженность нерестовых миграций весьма различна. Так, виды полупроходных карповых северной части Каспия (вобла, тарань) поднимаются вверх по рекам всего на несколько десятков километров. Громадные по протяженности миграции совершают многие лососевые. У дальневосточного лосося — кеты — миграционный путь достигает местами 2 тыс. км и более, а у сибирского вида — нерки — около 4 тыс. км. Семга поднимается по Печоре до ее верховьев. Несколько тысяч километров проходит на пути к местам нереста европейский речной угорь, размножающийся в западной части Атлантического океана, в Саргассовом море.
Вариабельно и время нерестовых миграций. У рыб оно вообще не может быть указано столь же определенно, как, например, сроки миграций птиц на гнездовье. Так, например, семга Белого моря входит в реки в два срока. Осенью идут особи со сравнительно слаборазвитыми половыми продуктами. Они зимуют в реке и размножаются на следующий год. Наряду с осенней есть и другая биологическая раса беломорской семги, которая входит в реки летом. Половые продукты у летних особей хорошо развиты, и они мечут икру в том же году. Такое поведение способствует экологической дифференцировке вида на две сезонные группы. У дальневосточной кеты также бывает два нерестовых хода. В Амур «летняя» кета заходит в июне — июле, «осенняя» — в августе — сентябре. В отличие от семги обе биологические расы кеты нерестуют в год захода в реку. Вобла заходит в реки каспийского бассейна для нереста весной, некоторые сиги в Сибири, наоборот, мигрируют к местам размножения только осенью.
Приведем обобщенные описания нерестовых миграций некоторых видов рыб.
Морская норвежская сельдь перед размножением нагуливается далеко к северо-западу от Скандинавии, у Фарерских островов, и даже в водах у Шпицбергена. В конце зимы косяки сельди начинают двигаться к берегам Норвегии и достигают их в феврале — марте. Нерест происходит в фьордах близ берега на мелких местах. Выведшиеся дичинки лишь отчасти остаются в фьордах, большая же их масса увлекается северо-восточной ветвью Гольфстрима — Нордкапским течением вдоль берегов Скандинавии на север. Такую пассивную миграцию личинки начинают зачастую в еще очень раннем возрасте, когда у них сохраняется желточный пузырь. За 3 — 4 месяца, до конца июля — начала августа, они проделывают путь в 1 000— 1200 км и достигают берегов Северной Норвегии (см. рис. 48).
Обратный путь молодые сельди проходят активно, но медленно — за 4 —5 лет. Они продвигаются на юг этапами ежегодно, то приближаясь к берегам, то отходя в открытое море. В четырех- или пятилетием возрасте сельди становятся половозрелыми и достигают к этому времени района икрометания — мест, где родились. Этим заканчивается первый, «юношеский», этап их жизни — период далекого путешествия на север. Второй период — период зрелости — связан с ежегодными миграциями от места нагула к местам нереста и обратно.
Нерестовые миграции проходных рыб рассмотрим на примере дальневосточных лососевых. Жизнь их в море в период полового созревания недостаточно изучена. Однако известно, что лососи держатся не распыленно по огромной акватории севера Тихого океана, а концентрируются в определенных его районах.
Ход кеты в реке Амур был хорошо изучен в свое время В. К. Солдатовым и его последователями. Как уже было сказано, там наблюдаются два ее хода: летний — с конца июня и осенний — в августе-сентябре.
Кета двигается из океана вверх по течению Амура со значительной скоростью — 30 — 35 км, а по некоторым данным — даже 47 км в сутки. Заходя в притоки Амура и двигаясь вверх по мелким речкам, рыба вынуждена преодолевать перекаты и пороги, где струи воды несутся с очень большой силой. Лососи вполне успешно перепрыгивают водопады высотой до 1 м. Перед препятствием они обычно отдыхают и набираются сил в местах со спокойной водой, затем быстро устремляются к водопаду и делают прыжок, вылетая из воды. При неудаче, отдохнув, рыбы вновь повторяют свою попытку.
Преодолевая препятствия, рыба, естественно, расходует огромное количество энергии. По П. Ю. Шмидту, расход энергии в сутки у кеты Равен у самцов 103 240 кДж, у самок — 113 560 кДж на 1 кг живой массы. При этом надо учесть, что, заходя в реки, рыбы перестают питаться и расход энергии происходит целиком за счет накопленных в теле питательных веществ. Это приводит к резким изменениям внешнего вида и физиологического состояния мигрантов.
По мере движения к местам нереста происходит созревание половых продуктов. Так, у кеты, проходящей в районе Амурского лимана, масса гонад самки составляет 7 % от общей массы тела рыбы; на расстоянии 300 км от устья реки масса гонад увеличивается до 10 % от массы тела, на расстоянии 570 км — до 12 %, на расстоянии 1 000 км — до 17 % и у места нереста (на расстоянии 11 90 км) — до 19%.
В местах нереста в притоках Амура, большей частью в их верховьях, в участках со спокойным течением, галечным грунтом и глубиной воды 0,5— 1,2 м кета появляется с развитыми половыми продуктами и вскоре начинает нереститься. Рыба откладывает икру в специально вырытые ямки и засыпает их галькой.
Проделав огромный и трудный путь, израсходовав почти всю энергию на дорогу и процесс размножения, рыбы, за весь этот период ничего не евшие, вскоре после нереста погибают: одни на местах нереста, другие после сноса течением рек вниз, в море. Берега рек бывают усеяны трупами кеты, здесь скапливается большое количество зверей и птиц, которые поедают ослабевшую и уже погибшую рыбу. Таким образом, кета размножается один раз в жизни. Мальки кеты весной следующего года начинают двигаться вниз по течению реки и летом достигают моря. Здесь они взрослеют, нагуливают вес, становятся половозрелыми и через 3 — 5 лет отправляются в странствие к местам, где они родились.
Чрезвычайно сложны и интересны связанные с размножением миграции европейского угря (Anguilla anguilla). Этот вид распространен по берегам Европы и Северной Африки, где обитает в самых разнообразных пресных водоемах — в реках, ручьях, прудах. Способность угрей расселяться очень велика, так как они могут переползать по суше из одного водоема в другой. Такие сухопутные миграции совершаются ночью, угри ползут по влажной траве иногда несколько километров.
В реках жизнь угрей начинается с появлением в их устьях небольших угревидных рыбок длиной 6 — 8 см и с совершенно прозрачным телом. Поднимаясь по рекам, угри темнеют и при длине около 20 см тело их покрывается чешуей. Продолжительность жизни их в реках различна, но во всяком случае не менее шести, а часто 10 лет и более.
Достигнув определенного возраста, угри приобретают брачный наряд и начинают мигрировать вниз по рекам в моря. Это явление внешне малозаметно, так как угри идут чаще ночью и по глубоким местам рек. По выходе в море угри становятся почти недоступными для наблюдений, поэтому долгое время значение этих миграций це было известно. Загадкой был и характер их размножения, так как у особей, живущих в реках, половые продукты незаметны. Это породило самые фантастические мнения. Только в конце XIX в. путем гистологических исследований удалось установить наличие у них семенников и яичников. Стало очевидным, что угри размножаются обычным половым путем. Но еще долгое время оставалось неясным, где же происходит их нерест. Ученые не знали ни зрелой икры угрей, ни их молоди.
Наряду со сказанным еще с начала прошлого столетия рыбакам и зоологам была известна маленькая прозрачная рыбка столь своеобразного строения, что ее выделили в особый род — Лептоцефалы (Leptocephalus). Эти рыбки различались размерами и строением; их было описано несколько видов. Только в 1897 г. опытным путем удалось установить, что лептоцефалы — это личинки угрей. Однако вопрос о месте их размножения оставался невыясненным.
Решить эту загадку удалось в начале XX в. знаменитому датскому ихтиологу Иоганну Шмидту. В схеме жизненный цикл угрей представляется так. Взрослые угри выходят из рек Европы и Северной Африки и, двигаясь на запад или юго-запад, достигают западных частей Атлантического океана. Они двигаются со средней скоростью 15 км в сутки, иногда их скорость достигает 40 — 50 км в сутки. Их путь равен 7 — 8 тыс. км. Весной, достигнув области между Бермудскими и Багамскими островами, угри опускаются на глубину около 1000 м, нерестятся и погибают. Выведшиеся личинки поднимаются из глубин, куда не проникает ни один луч света, на залитую тропическим солнцем поверхность океана и начинают обратное странствие на восток через весь океан. Они двигаются в значительной мере пассивно, влекомые струями Гольфстрима. Три года продолжается это путешествие, и вот у берегов Европы и Африки появляются прозрачные угри, которые входят в реки, где начинается вторая фаза их жизни. Через 6 —9 (а иногда и через 10 — 20) лет, уже взрослыми, они вновь направляются в океан, где оставляют потомство, а сами погибают.
Как ориентируются рыбы, отыскивая места нереста, отстоящие от мест нагула на тысячи километров? Как и в связи с чем возникли явления нерестовых миграций? В общем виде дать ответ на первый вопрос сравнительно легко.
Несомненно, что в основе миграционных движений рыб лежит их нервно-чувствующая рефлекторная деятельность. При миграциях рыбы воспринимают закономерные и постепенные изменения Условий среды, играющие роль своего рода сигналов-раздражителей. Так, проходные лососи ориентируются, видимо, на постепенные изменения химизма воды. Мигрирующие угри также руководствуются, видимо, закономерными изменениями температуры и солености воды. Дело в том, что место их нереста является областью самой теплой и соленой воды в океане.
Вышедшие из любого пункта европейского или африканского побережья угри, если они будут ориентироваться на постоянно повышающуюся температуру и соленость воды, должны будут прийти кратчайшим путем к области наиболее теплой и соленой воды, т.е. к месту своего икрометания.
Руководит рыбами при их тысячеверстных странствиях удивительная по тонкости способность воспринимать особенности химического и физического состояния воды и связанная с этим феноменальная память на соответствующие раздражения.
В пользу доминирующего значения обонятельной ориентации рыб при миграциях говорят и экспериментальные данные: лососи с изолированными ноздрями теряли способность к отысканию верного пути и мигрировали в разных, несвойственных им направлениях.
Причины миграции угрей еще не получили должного объяснения. Согласно данным П. Ю. Шмидта, во время максимального оледенения распределение температур в Атлантике было иным и область наиболее теплой воды располагалась далеко к югу от современного не положения и имела вид не овала ограниченных размеров в западных частях океана, как в настоящее время, а составляла более или менее широкую полосу, проходившую через весь океан на восток. Миграционный путь угрей, как европейских, так и американских, был в этой связи коротким.
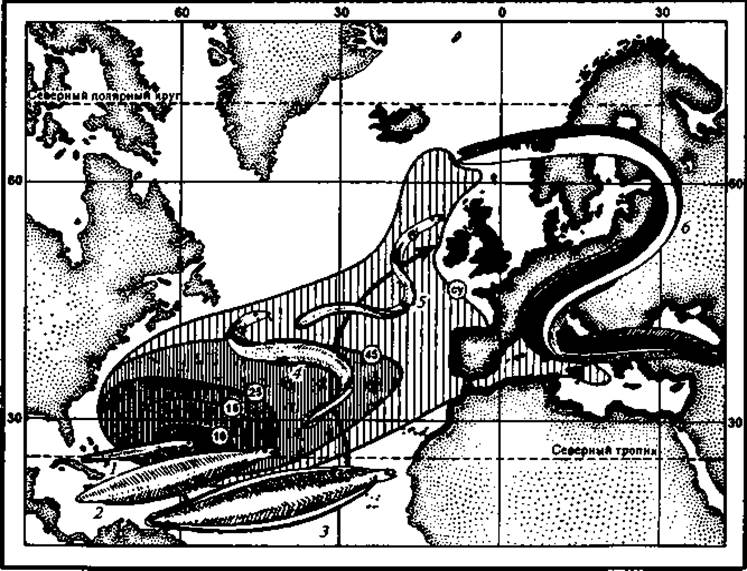
Рис. 49. Схема распространения и миграций европейского угря:
1 — только что вышедшая личинка; 2 — годовалая личинка; 3 — двухгодовалая личинка; 4 — личинка перед началом превращения; 5 — стеклянный угорь; 6 — взрослый угорь; 7— ареал стадии стеклянного угря; 8 — области распространения европейского угря в пресных водоемах материков; 9 — средняя длина личинок, мм
С окончанием ледникового периода, когда Гольфстрим принял свое современное северо-восточное направление и, с другой стороны, в восточную часть океана проникли холодные воды Южного полушария, область максимальных температур сократилась и отодвинулась к западу. Угри по мере отодвигания ледника и появления на материках новых рек заселяли их. Расселение угрей на север и сокращение и отодвигание области наиболее теплой воды на запад и вызвали удлинение миграционных путей (рис. 49).
На миграции проходных лососей существуют два противоположных взгляда. То обстоятельство, что почти все проходные рыбы распространены в Северном полушарии, делает весьма вероятной также зависимость возникновения миграций от явлений ледникового периода.
По одному из предположений, опреснение морских вод в период таяния ледников сделало возможным выход взрослых речных рыб в моря, более богатые кормом, чем реки. В реки рыбы возвращались для размножения. Условия для этого здесь более благоприятны: меньше хищников и конкурентов, лучше обеспеченность воды кислородом и др.
По другой гипотезе, проходные рыбы были исконно морскими и их заход в реки — явление вторичное, связанное с сильным опреснением морей во время таяния ледников, что, в свою очередь, позволило рыбам легче приспособиться к жизни в пресной воде. Несомненно, что проходные лососевые меняют места своего обитания в зависимости от биологического состояния. Взрослые рыбы населяют обширные пространства морей, богатые кормом. Их молодь выводится в тесных пресных водоемах (верховья рек). Здесь невозможно существование всей массы выросших рыб в силу ограниченности пространства и недостатка корма, однако для вывода молоди условия более благоприятны. Это обусловливается чистой, богатой кислородом водой, возможностью закапывания икры в грунт Дна и ее успешным развитием в пористом грунте.
Природа кормовых миграций вполне понятна, если учесть, что в период нереста рыбы выбирают условия среды, которые, как правило, не представляют большой ценности в кормовом отношении. Кроме того, большинство рыб во время размножения перестают питаться, и, следовательно, после нереста потребность в корме резко возрастает. Это заставляет рыб искать области с особо благоприятными кормовыми возможностями. Примеров кормовых миграций очень много. Рассмотрим некоторые из них.
Европейский лосось — семга в отличие от своего тихоокеанского сородича — кеты после нереста не погибает, и передвижения отнерестившихся рыб вниз по реке должны рассматриваться как кормовые миграции. Но и после выхода в море рыбы совершают закономерные массовые миграции в поисках мест, особо богатых пищей. Каспийская севрюга, вышедшая после нереста из Куры, пересекает Каспийское море и нагуливается преимущественно у его восточного побережья.
Широко мигрирует в поисках пищи атлантическая треска. Одним из основных мест ее нереста служат отмели (банки) у Лофотенских островов в Атлантическом океане. После размножения треска становится чрезвычайно прожорливой, и в поисках пищи большие ее стаи направляются вдоль берегов Скандинавии на северовосток и далее на восток через Баренцево море к острову Колгуеву и Новой Земле, частью на север, к Медвежьему острову и далее к Шпицбергену (рис. 50). Эта миграция представляет для нас особый интерес, так как промышленный лов трески в районе Мурманска и в Канинско-Колгуевском мелководье в значительной мере базируется на добыче мигрирующих и кормящихся косяков. Треска при миграциях придерживается теплых струй Нордкапского течения, по которым проникает через Карские Ворота и Югорский Шар даже Карское море. Наибольшее количество трески в Баренцевом море скапливается в августе.
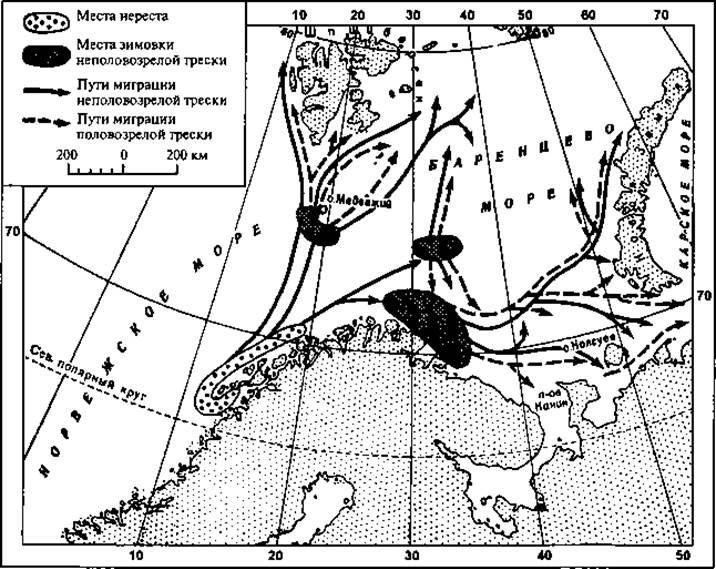
Рис. 50. Схема миграций взрослой трески
Многие виды рыб при зимнем понижении температуры воды становятся малоактивными или даже впадают в состояние оцепенела. В этом случае они не остаются обычно в местах нагула, а собираются на ограниченных пространствах, где условия рельефа дна, фунта и температуры благоприятствуют зимовке. Это зимовальные миграции. Так, сазан, лещ, судак мигрируют в низовья Волги, Урала, Куры и других больших рек, где, скапливаясь в огромном количестве, залегают в ямы. Издавна известна зимовка осетров в ямах на реке Урал. В состоянии спячки рыбы лежат неподвижно. У многих видов поверхность тела в это время покрывается толстым слоем слизи, которая в известной мере изолирует рыб от отрицательного действия низких температур. Обмен веществ у зимующих таким образом рыб сильно понижается. Некоторые рыбы, например караси, зимуют, закапываясь в ил. Известны случаи, когда они вмерзают в ил и успешно перезимовывают, если «соки» их тела не подвергаются замораживанию. Опыты показали, что лед может окружать все тело рыбы, но внутренние «соки» остаются незамерзшими и имеют температуру до 0,2—0,3 °С.
Не всегда зимовальные миграции заканчиваются впаданием рыб в состояние оцепенения. Так, азовская хамса по окончании нагула на зиму выходит из Азовского моря в Черное. Это обусловлено неблагоприятными условиями, возникающими зимой в Азовском море в связи с появлением ледяного покрова и сильным охлаждением этого неглубокого водоема.
Питание. Характер пищи у рыб чрезвычайно разнообразен. Рыбы кормятся почти всеми живыми существами, обитающими в воде, от мельчайших планктонных растительных и животных организмов до крупных позвоночных. При этом сравнительно немногие виды кормятся только растительной пищей, большинство же поедает животных или смешанную животно-растительную пищу. Деление рыб на хищных и мирных в значительной мере условно, так как характер пищи существенно варьирует в зависимости от условий водоема, времени года и возраста рыбы.
Особо специализированными растительноядными видами являются планктоноядные толстолобы (Hypopthalmichthys) и поедатели высшей растительности белые амуры (Ctenopharyngodon) из отряда Карпообразные.
Преимущественно растительноядны краснопёрка (Scardinius), маринка (Schizothorax) и храмуля (Varicorhins). Большинство рыб кормится смешанной пищей. Однако в молодом возрасте все рыбы проходят стадию мирного питания планктоном и по мере взросления переходят на свойственную им пищу (бентос, нектон, планктон). У хищников переход на рыбный стол происходит в различном возрасте. Так, щука начинает заглатывать личинок рыб, достигнув длины тела всего 25 — 33 мм, судак — 33 — 35 мм; окунь же переходит на рыбное питание сравнительно поздно, при длине тела в 50—150 мм, при этом главную пищу окуня в течение 2 — 3 первых лет его жизни составляют беспозвоночные.
Разнообразны приемы добывания пищи. Многие хищники преследуют свою жертву, догоняя ее в открытой воде. Таковы акулы, жерех, судак. Есть хищники, подкарауливающие добычу и хватающие ее накоротке. Так охотятся, например, щуки, сомы. Пила-рыба и пилонос используют при охоте свой мечевидный орган. С большой скоростью врезаются они в стаи рыб и делают мечом несколько сильных ударов, которыми убивают или оглушают жертву. Насекомоядная рыба полосатый брызгун (Toxotes jaculator) имеет специальное приспособление, посредством которого она выбрасывает сильную струю воды, сбивающую насекомых с прибрежной растительности.
Многие донные рыбы раскапывают грунт и выбирают из него кормовые объекты. Карп способен доставать пищу, проникая в толщу грунта на глубину до 15 см, лещ — только до 5 см, окунь же берет корм с его поверхности. Успешно копаются в грунте американские веслоносы (Polyodon) и среднеазиатские лжелопатоносы (Pseudoscaphirhynchus), использующие для этого свой рострум (обе рыбы из отряда Осетрообразные).
Крайне своеобразно приспособление для добывания пищи у электрического угря (Electrophorus electricus). Эта рыба, перед тем как схватить свою добычу, поражает ее электрическим разрядом, достигающим у крупных особей 300 В.
Интенсивность питания рыб в течение года и жизненного цикла неодинакова. В целом в период нереста подавляющее большинство видов перестают кормиться, сильно худеют. Так, у атлантического лосося масса мышц уменьшается более чем на 30 %. Поэтому потребность в пище у этих рыб в другие этапы жизненного цикла исключительно велика. Посленерестовый период носит название периода восстановительного питания или «жора».
Размножение. Подавляющее большинство рыб раздельнополы. «Рыбий стиль» размножения характеризуется наружным оплодотворением, многочисленными половыми продуктами, как правило, отсутствием заботы о потомстве. Размер гонад у рыб ко времени размножения резко увеличивается. Нередки случаи, когда масса гонад в это время равна 25 % и даже более от общей массы тела.
Сравнительно с другими позвоночными рыбы характеризуются громадной плодовитостью. Достаточно указать, что большинство видов в год откладывают сотни тысяч икринок, а, например, треска — до 10 млн, луна-рыба — даже сотни миллионов икринок. Громадная плодовитость рыб понятна, если учесть, что яйца у подавляющего большинства видов оплодотворяются вне тела матери, когда вероятность оплодотворения резко снижается. К тому же спермин сохраняют в воде способность к оплодотворению очень недолго, в течение короткого времени, хотя и различного в зависимости от условий нереста. Так, у кеты и горбуши, нерестящихся на быстром течении, где контакт спермы с икрой может осуществляться в очень малый промежуток времени, спермин сохраняют подвижность лишь на протяжении 10—15 с, у русского осетра и севрюги, нерестящихся на более медленном течении, — 230 — 290 с. У волжской сельди через минуту после выделения спермы в воду подвижность сохраняло только 10 % спермиев, а через 10 мин двигались лишь единичные сперматозоиды, у видов, нерестящихся в относительно малоподвижной воде, спермин сохраняют подвижность дольше. Так, у океанической сельди спермин сохраняют способность к оплодотворению более суток.
Икринки, попадая в воду, продуцируют стекловидную оболочку, которая вскоре не дает возможности спермиям проникнуть внутрь. Все это уменьшает вероятность оплодотворения. Опытные подсчеты показали, что у лососей Дальнего Востока процент оплодотворенной икры равен 80. У некоторых рыб этот процент еще меньше.
Развитие зародышей происходит в воде, они ничем не защищены и не охраняются. В силу этого вероятность гибели развивающихся личинок и мальков рыб очень велика. Для промысловых рыб Северного Каспия установлено, что из всех выведшихся из икры личинок не больше 10 % скатывается в море в виде сформировавшихся рыбок, остальные 90 % погибают.
Очень мал процент рыб, доживающих до половозрелости. Например, для севрюги он определяется в 0,01 %, для осенней кеты Амура — 0,13 — 0,58 %, для атлантического лосося — 0,125 %, для леща — 0,006 — 0,022 %.
Таким образом, очевидно, что громадная начальная плодовитость рыб служит важным биологическим приспособлением для сохранения видов. Справедливость этого положения доказывается еще и ясной зависимостью между плодовитостью и условиями размножения.
Наибольшей плодовитостью (миллионы икринок) отличаются морские пелагические рыбы и виды, имеющие плавающую икру. Вероятность гибели последней особенно велика, так как она легко может быть съедена другими рыбами, выброшена на берег и т. п. Рыбы, откладывающие тяжелую, оседающую на дно икру, которая к тому же обычно приклеивается к водорослям или камням, менее плодовиты. Многие лососевые откладывают икру в специально сооружаемые ими ямы, а некоторые засыпают потом эти ямы мелкой галькой. В этих случаях как бы проявляются первые признаки заботы 0 потомстве. Соответственно с этим уменьшается и плодовитость. Так, семга выметывает от 6 до 20 тыс. икринок, кета — 2 — 5 тыс., а горбуша — 1 — 2 тыс. Укажем для сравнения, что севрюга откладывает до 400 тыс. икринок, осетры — 400 тыс. —2 млн 500 тыс., белуга — 300 тыс.—8 млн, судак — 300 — 900 тыс., сазан — 400 тыс.—1 млн 500 тыс., треска — 2 млн 500 тыс. — 10 млн. Эти виды такой заботы о потомстве не проявляют.
Наконец, большинство хрящевых рыб, имеющих внутреннее осеменение и яйца с плотной оболочкой (которые они укрепляют на камнях или водорослях), откладывают их единицами или десятками.
С возрастом у большинства рыб плодовитость повышается и лишь к старости несколько снижается. Следует иметь в виду, что большинство промысловых рыб не доживают до старости, так как к этому времени они бывают уже выловлены.
Как указано ранее, для огромного большинства рыб характерно наружное оплодотворение. Исключение составляют почти все современные хрящевые рыбы и некоторые костистые. У первых в качестве копулятивного органа функционируют крайние внутренние лучи брюшных плавников, которые самцы при спаривании складывают вместе и вводят в клоаку самки. Много видов с внутренним оплодотворением в отряде Карпозубообразные (Cyprinodontiformes). Копуля- тивным органом у этих рыб служат видоизмененные лучи анального плавника. Внутреннее оплодотворение характерно для морского окуня (Sebastes marinus), хотя копулятивных органов у него нет.
В отличие от большинства позвоночных рыбы (если говорить о надклассе вообще) не имеют определенного сезона размножения. По времени нереста можно выделить по крайней мере три группы рыб.
Первую составляют нерестящиеся весной и ранним летом — осетровые, карповые, сомовые, сельди, щуки, окуни и др.
К нерестящимся осенью и зимой принадлежат главным образом рыбы северного происхождения. Так, атлантический лосось семга начинает нереститься с начала сентября; период икрометания растягивается у него в зависимости от возраста рыбы и условий водоема до конца ноября. Поздней осенью нерестится речная форель. Сиги мечут икру в сентябре—ноябре. Из морских рыб треска нерестится у побережья Норвегии с декабря по июнь, а у берегов Кольского полуострова — с января до конца июня.
Как указывалось ранее (см. раздел «Миграции»), у проходных рыб, например у кеты и семги, имеются биологические расы, различающиеся временем захода в реки для нереста.
Существует, наконец, третья группа рыб, которые вовсе не имеют определенного срока размножения. К ним принадлежат главным образом тропические виды, температурные условия обитания которых в течение года существенно не меняются. Таковы, например, виды семейства Цихлидовые (Cichlidae).
Места нереста крайне разнообразны. В море рыбы откладывают икру, начиная от зоны приливов и отливов, например пинагоры (Cyclopterus), атерина (Atherina) и ряд других, и до глубин 500—1 000 м, где нерестятся угри, некоторые камбалы и др. Треска и морские сельди мечут икру у берегов, на сравнительно мелких местах (банках), но уже вне зоны приливов и отливов.
Не менее разнообразны условия нереста в реках. Лещ в низовых ильменях Волги откладывает икру на водных растениях. Жерех, наоборот, выбирает места с каменистым дном и быстрым течением. В заводях, заросших водорослями, нерестятся окуни, которые прикрепляют икру к подводной растительности. На очень мелких местах, заходя в небольшие речки и канавы, нерестятся щуки.
Условия, в которых находится икра после оплодотворения, весьма разнообразны. Большинство рыб оставляют ее на произвол судьбы, но некоторые помещают икру в специальные сооружения и более или менее длительное время ее охраняют. Бывают, наконец, случаи, когда рыбы носят оплодотворенную икру на своем теле или даже внутри своего организма.
Забота о потомстве. Приведем некоторые примеры заботы рыб о потомстве. Нерестилища кеты расположены в мелких притоках Амура, в местах с галечным грунтом и относительно спокойным течением, глубиной 0,5 — 1,2 м; при этом важно наличие подземных ключей, дающих чистую воду. Самка, сопровождаемая одним или несколькими самцами, найдя место, пригодное для откладывания икры, ложится на дно и, судорожно изгибаясь, расчищает его от травы и ила, поднимая при этом облако мути. Далее самка также ударами хвоста и изгибанием всего тела вырывает в грунте яму. После этого начинается сам процесс нереста. Самка, находясь в яме, выметывает икру, а самец, находящийся рядом с ней, выпускает молоки. Около ямы обычно стоят несколько самцов, между которыми часто бывают драки.
Икра откладывается в яме гнездами, которых обычно бывает три. Каждое гнездо засыпается галькой, а когда закончено сооружение последнего гнезда, самка насыпает над ямой бугор овальной формы (длиной 2 — 3 м и шириной 1,5 м), который сторожит несколько дней, мешая другим самкам вырыть здесь яму для икрометания. Вслед за этим самка погибает. Сходно с кетой строит гнездо нерка (рис. 51).
Еще более сложное гнездо делает трехиглая колюшка. Самец вырывает на дне ямку, выстилает ее обрывками водорослей, затем устраивает боковые стенки и свод, склеивая растительные остатки клейким выделением кожных желез. В готовом виде гнездо имеет форму шара с двумя отверстиями. Затем самец загоняет в гнездо одну за другой нескольких самок и поливает каждую порцию икры (по 20— 100 икринок) молоками, после чего в течение 10 — 15 дней охраняет гнездо от врагов. При этом самец располагается относительно гнезда таким образом, что движения его грудных плавников возбуждают ток воды, идущий над икрой. Этим обеспечивается лучшая аэрация, а следовательно, и более успешное развитие икры.
Дальнейшие усложнения заботы о потомстве можно видеть у рыб, Которые носят оплодотворенную икру на своем теле.
У самки сомика аспредо (Aspredo filamentosus) кожа на брюхе в период нереста заметно утолщается и размягчается. После выметывания икры и оплодотворения ее самцом самка тяжестью своего тела вдавливает икру в кожу брюха. Кожа приобретает вид мелких сотов, в ячейках которых сидят икринки. Последние связаны с телом матери развивающимися стебельками, снабженными кровеносными сосудами (рис. 52). У самцов иглы-рыбы (Syngnathus acus) и морского конька (Hippocampus) на нижней стороне тела имеются кожистые складки, образующие своего рода яйцевой мешок, в который самки откладывают икру. У морской иглы (рис. 53) складки лишь загибаются на брюхо и прикрывают икру. У морского конька приспособление к вынашиванию развито еще сильнее. Края яйцевого мешка у него плотно срастаются, на внутренней поверхности образовавшейся камеры развивается густая сеть кровеносных сосудов, через посредство которых, видимо, и осуществляется газообмен эмбрионов.
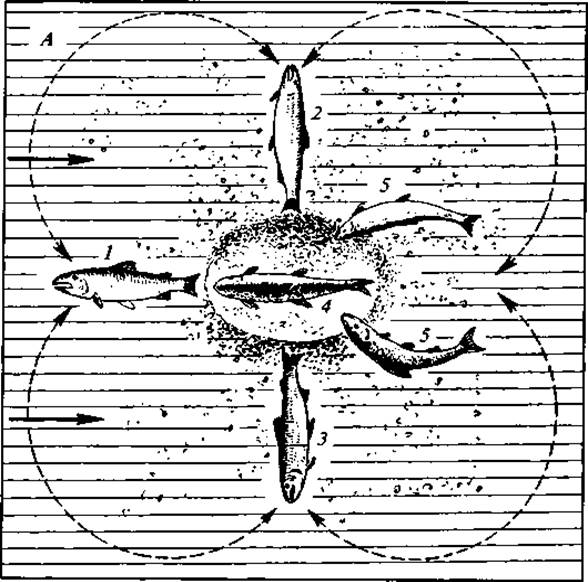

Рис. 51. Постройка гнезда и икрометание у нерки:
А — вид сверху; Б — в разрезе; 1—3 — положение самки при выкапывании грунта 4 — главный самец; 5 — второстепенные самцы

Рис. 52. Самка сомика аспредо с икрой
Есть виды, вынашивающие икру во рту.
Так бывает у американского морского сома (Galeichthys felis), у которого самец в ротовой полости вынашивает до 50 яиц, и у кардиналок (рис. 54). В это время он, видимо, не питается. У других видов (например, рода Tilapia) икру носит во рту самка. Иногда во рту бывает более 100 икринок, которые самка осторожно передвигает, что связано, видимо, с обеспечением лучшей аэрации. Инкубационный период (судя по наблюдению в аквариуме) длится 10—15 дней. В это время самки почти не питаются. Любопытно, что и после вывода мальки еще некоторое время при опасности прячутся в рот матери.
Упомянем о своеобразном размножении горчака (Rhodeus sericeus) из семейства карповых, широко распространенного на юге России и в Украине. В период нереста у самки развивается длинный яйцеклад, которым она откладывает икру в мантийную полость моллюсков (Unio или Anodonta, рис. 55). Самец выделяет молоки, находясь рядом с моллюском. Яйца оплодотворяются сперматозоидами, всасываемыми моллюском с током воды через сифон. Эмбрионы развиваются на жабрах моллюска и выходят в воду, достигнув длины около 10 мм. Последняя степень усложнения процесса размножения у рыб выражается в живорождении. Икра не поступает в наружную среду, и личинки развиваются в половых путях самки. Развитие осуществляется отчасти за счет желтка яйца, отчасти за счет материнского организма. Последнее обеспечивается тесным контактом (но не слиянием) густой сети кровеносных сосудов желточного мешка эмбриона с кровеносными сосудами слизистой матки родителя. Функционально (но не анатомически) это образование подобно детскому месту млекопитающих и обозначается как желточная плацента.
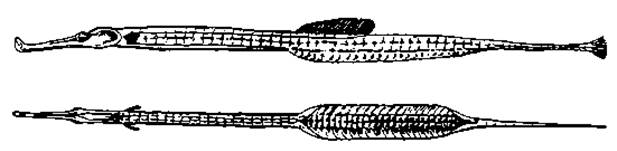
Рис. 53. Самец морской иглы с яйцевым мешком. На нижнем рисунке мешок раскрыт и видна икра

Рис. 54. Самец кардиналки (Apogon imberbis) с икрой в ротовой полости
Ведущее значение желточной плаценты в газообменных процессах и в удалении из организма эмбриона азотистых продуктов метаболизма дает основание говорить о настоящем живорождении.
Живорождение в наибольшей мере характерно для хрящевых рыб, у которых оно наблюдается даже чаще, чем откладывание яиц. Наоборот, среди костных рыб это явление встречается очень редко. В качестве примера можно указать на байкальских голомянок (Соmephoridae), морских собачек (Blenniidae), морских окуней (Serranidae) и особенно карпозубых (Cyprinodontidae). Все яйцеживородящие рыбы имеют малую плодовитость. Большинство рождают единицы детенышей, реже десятки.
Мы привели ряд случаев, когда рыбы не оставляют оплодотворенную икру на произвол судьбы и проявляют в том или ином виде заботу о ней и о развивающейся молоди. Такая забота свойственна ничтожному меньшинству видов. Основным, в наибольшей мере характерным типом размножения рыб является такой, при котором икра оплодотворяется вне тела матери и в последующем родители оставляют ее на произвол судьбы. Именно этим и объясняется огромная плодовитость рыб, обеспечивающая сохранение видов при очень большой, неизбежной в указанных условиях, гибели икры и молоди.
Подавляющее число видов рыб раздельнополы, для них характерен «рыбий стиль» размножения. Исключение составляют немногий костные рыбы, например морские окуни (Serranus scriba) и некоторые другие. Им свойствен гермафродитизм. Как правило, в случае гермафродитизма половые железы функционируют попеременно то как семенники, то как яичники, и самооплодотворение в силу этого невозможно. Только у морского окуня разные части гонады одновременно выделяют яйца и сперматозоиды. Иногда встречаются гермафродитные особи у трески, макрели, сельдей.
У некоторых рыб иногда наблюдается партеногенетическое развитие, которое, однако, не приводит к формированию нормальной дичинки. У салаки и тихоокеанской сельди партеногенетическое развитие доходит до стадии свободноплавающей личинки. Есть и другие примеры подобного рода.
У лососей неоплодотворенные икринки, заложенные в гнезде, не погибают и своеобразно развиваются (происходит деление клеток) до того времени, когда из оплодотворенных икринок выводятся мальки. Это своеобразное приспособление к предохранению кладки.
Известен у рыб и другой тип отклонения от нормального размножения, именуемый гиногенезом. В этом случае спермин проникают в яйцо, но слияния ядер яйцеклетки и спермия не происходит. Развитие идет нормально, но в потомстве развиваются только одни самки. Так бывает у серебряного карася и прослеживается географически. В Восточной Азии обнаруживаются как самки, так и самцы этого вида, и размножение протекает нормально. В Средней Азии, Западной Сибири и Европе самцы встречаются крайне редко, а в некоторых популяциях их нет вовсе. В таких случаях осеменение, приводящее к гиногенезу, осуществляется самцами других видов рыб (Г. В. Никольский, 1961).
Рост и возраст. Продолжительность жизни рыб различна. Есть виды, живущие немногим более года: некоторые бычки (Gobiidae) и светящиеся анчоусы (Scopelidae). С другой стороны, белуга доживает до 100 лет и более. Некоторые камбалы живут 50 — 60 лет. Вo всех этих случаях имеется в виду предельная потенциальная продолжительность жизни. В условиях же регулярного промысла действительная продолжительность жизни много меньше (табл. 5).

Рис. 55. Горчак, откладывающий икру
Таблица 5
Предельные размеры и возраст некоторых видов рыб
(по Г. В. Никольскому, 1961)
Вид рыб |
Водоем |
Предельные размеры, см |
Возраст, годы |
Хамса |
Азовское море |
13 |
3 |
Шпрот |
Балтийское море |
16 |
6 |
Вобла |
Северный Каспий |
35 |
10 |
Сазан |
Амур |
90 |
16 |
Океаническая сельдь |
Норвежское и Северное моря |
37 |
23 |
Треска |
Баренцево море |
169 |
25 |
Севрюга |
Кура |
214 |
31 |
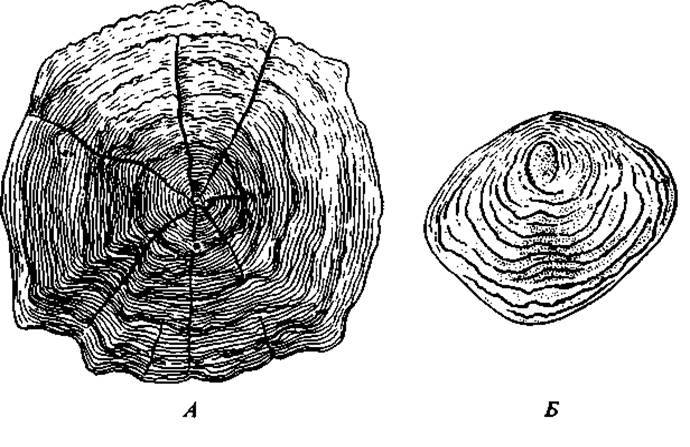
Рис. 56. Чешуи с годичными кольцами у разных рыб: А — вобла; Б — треска
Рост рыбы в отличие от большинства позвоночных, как правило, не прекращается по достижении половой зрелости, а продолжается до старости. Отметим, что для рыб характерна четко выраженная сезонная периодичность роста. Летом, особенно во время нагула, они растут значительно быстрее, чем в малокормный зимний период. Эта неравномерность роста сказывается на строении ряда костей и чешуи. Периоды замедленного роста соответствуют узким полосам или кольцам. При рассмотрении в падающем свете они кажутся светлыми, в проходящем свете, наоборот, темными. В периоды усиленного роста откладываются широкие кольца или слои, которые в проходящем свете кажутся светлыми. Комбинация двух колец — узкого зимнего и широкого летнего — и представляет годовую отметку. Подсчет этих отметок позволяет определить возраст рыбы (рис. 56).
Определение возраста рыб имеет огромное теоретическое и практическое значение. При рационально поставленном промысле анализ возрастного состава улова служит важнейшим критерием для установления перелова или недолова. Увеличение в улове долевого значения младших возрастов и уменьшение старших указывает на напряженность промысла и угрозу перелова. Наоборот, большой процент особей старших возрастов говорит о неполном использовании запасов рыбы.
Практическое значение рыб
Значение рыб в жизни человечества исключительно велико. В белковом рационе человека в разных странах мира рыба составляет от 17 до 83 %. Кроме пищевых продуктов из рыбы получают витамины, кормовую муку, удобрения и др.
Рыба — основной извлекаемый человечеством из водной среды биологический продукт. Ее значение в общем вылове (по массе) равно примерно 85 %. Основную часть рыбы (около 90 %) добывают в морях. Из трофических групп наибольшее значение имеют планктонофаги — 65 %, 25 % хищные и около 10 % бентософаги.
Проблема рыбного промысла и хозяйствования в связи с этим Промыслом может быть охарактеризована рядом ее форм (морской Промысел, добыча в пресных водоемах, разведение, акклиматизация).
Морской промысел. Общая площадь Мирового океана исключительно велика, при этом площадь океанов с глубинами более 3 тыс. м составляет 50 —60 %. Рыбы обнаружены на глубинах до 10 тыс.м. Все это, казалось бы, рисует Мировой океан как плацдарм для неограниченного увеличения добычи морских продуктов, в частности и рыбы. Однако, хотя современная техника рыбного промысла позволяет вылавливать рыб, живущих на больших глубинах, опыт рыбопромысловой практики показывает, что наибольшее сосредоточение рыб и самые легкие возможности их добывания имеются на ничтожной части Мирового океана. Это прибрежная зона, которая характеризуется малыми глубинами (до 200 м) и известна под названием шельфа, или материковой ступени. Суммарная площадь шельфов всех океанов и сопутствующих им морей равна 8 % всей площади Мирового океана. Но именно в области шельфа добывается 90 % всего мирового улова рыбы.
Во второй половине XX в. вылов рыбы в Мировом океане в хозяйствовании России резко увеличился. Рыбу добывают теперь не только в таких окраинных морях, как Баренцево, Берингово, Охотское. Японское, но и в открытых пространствах Атлантического, Индийского и Тихого океанов.
По отдельным океанам и сопутствующим им морям долевое значение улова рыбы выглядит примерно так: Тихий и Атлантически і і океаны — по 40 — 45%, Индийский океан — около 10%, Северный Ледовитый океан — всего около 5 %. Размещение общей добычи по широтным зонам может быть иллюстрировано такими цифрами: северная зона (Северный Ледовитый океан и северные части Атлантического и Тихого океанов с сопутствующими морями) — примерно 40—50 %, тропическая зона (соответствующие части Атлантического. Тихого и Индийского океанов) — около 30 %, Южное полушарие — около 10%. Рост вылова в указанных зонах был также весьма неравномерным: в северной зоне он увеличился во второй половине XX в. примерно на 50 %, в тропической зоне — в 4 раза, а в Южном полушарии — более чем в 10 раз. В обиход рыбного стола жителей России было введено много незнакомых раньше видов рыб: хек. минтай, сквама, сабля, нототения, ледяная и многие другие.
Добыча рыбы на шельфах осложняется внедрением в эти зоны других форм хозяйствования, в частности добычи нефти, газа. Шельфовая зона страдает от обильного курсирования танкеров и аварийного изливания их содержимого. Серьезную проблему создает возникающий временами перелов отдельных видов (морской окунь, треска, нототения и пр.).
Заключение в 1970-х годах Всемирного соглашения о 200-мильной охранной зоне усложнило для России добычу рыбы в Атлантическом и Тихом океанах.
Добыча рыбы в пресных водоемах. В первой трети XX столетия в России основной вылов рыбы производился во внутренних водоемах: в первую очередь в районах Северного Каспия и низовьях впадающих в него рек, Черного моря, в реках Сибири и Дальнего Востока. Это касалось таких проходных групп рыб, как осетровых и лососевые.
Гидростроительство на Волге, Дону, Днепре и других реках создало серьезные проблемы для вылова этих ценных рыб: преграды на путях миграций рыб к местам нереста, возникновение огромных водохранилищ. Плотины преграждают миграционные пути ценных рыб (лососевых и осетровых), естественные нерестилища которых расположены значительно выше (по течению рек) плотин. В связи с этим возникает проблема строительства рыбопропускных сооружений. Последние могут быть двух типов: лестницы-рыбоходы (рис. 57) и рыбоподъемники.
Лестница-рыбоход, устроенная в районе Туломской гидроэлектростанции, состоит из 57 ступеней с перепадами между ними по 30 см. По этой водной лестнице из нижнего бьефа в верхний на высоту 16—19 м поднимаются для нереста семга, кумжа, хариус. Скорость течения воды на порогах ступенек лестницы достигает 2,5 м/с, что не представляет для лососевых рыб существенных препятствий, так как и в естественных условиях им при ходе на нерест приходится преодолевать большие быстрины и водопады.
В Канаде при сооружении лестниц-рыбоходов устраивают и камеры для отдыха поднимающейся по ступенькам рыбы. Камеры отдыха сооружают на расстоянии 300 м друг от друга.
Пропуск через плотину осетровых рыб не может быть обеспечен сооружением подобных лестниц-рыбоходов, так как эти рыбы плохо преодолевают препятствия в виде водопадов. Здесь необходима разработка иных мероприятий. Наблюдения показали, что многие особи проходных осетровых в таких случаях размножаются не доходя до первой плотины на своем миграционном пути. Значение этой естественной черты может быть повышено путем искусственного создания участков дна с оптимальными для нереста условиями.
Важное значение имеет искусственное разведение ценных проходных рыб.
Искусственное разведение ценных проходных рыб основано на том, что при размножении в естественных условиях далеко не вся икра оказывается оплодотворенной, огромный процент икры и мальков погибает. Так, например, на Амуре во время нереста «осенней» кеты потеря икры составляет в среднем 20 — 40%. Много мальков гибнет в первые периоды их жизни: ко времени ухода с нерестилищ в море в живых остается лишь около 10 % мальков кеты. Много мальков гибнет по пути с нерестилищ в море, так как зачастую путь их равен 1 — 1,5 тыс. км. В итоге моря достигает ничтожное количество мальков, иногда менее 1 %. Естественно, что с такими непроизводительными потерями нельзя мириться.

Рис. 57. Лестница-рыбоход на реке Тулома
Еще в начале текущего столетия известный знаток дальневосточных лососевых В. К. Солдатов настойчиво рекомендовал искусственное разведение этих ценных рыб.
Искусственное разведение проходных рыб осуществляется в специальных аппаратах. Широкое распространение в России нашел так называемый «сухой» способ оплодотворения икры, предложенный более 100 лет назад русским рыбоводом В. П. Врасским. Он основан на том, что в воде спермин быстро теряют подвижность и часть икры остается неоплодотворенной. При применении метода Врасского икру выпускают в таз, в который затем отцеживают молоки самцов. Содержимое таза осторожно перемешивают так, чтобы молоки равномерно распределились среди икринок и вероятность оплодотворения стала наибольшей. Только после этого в таз наливают воду.
Оплодотворенную икру закладывают в аппараты с проточной водой, где она развивается при постоянном контроле специалистами. Мертвые и заболевшие икринки регулярно удаляют. Период инкубации икры кеты длится 103 — 120 дней. Выведшихся мальков содержат в специальных, отгороженных от реки, водоемах, где питание мальков осуществляется эндогенно, за счет энергетических веществ желточного мешка. Последний рассасывается только через 60 — 90 дней после вылупления эмбрионов из яйца (рис. 58). В последующем мальков, переходящих на экзогенное питание, выпускают в реку, по которой они скатываются в море.
Результаты работы рыборазводных заводов весьма показательны. Так, на заводе, разводящем кету, оплодотворение икры составляет 98 —99 %. Гибель икры во время инкубации составляет всего 4—8 %, а гибель личинок не превышает обычно 0,5 %. Общее количество скатывающейся молоди составляет более 90 % от инкубируемой икры, в то время как при естественном размножении лососей этот процент равен всего лишь 10.
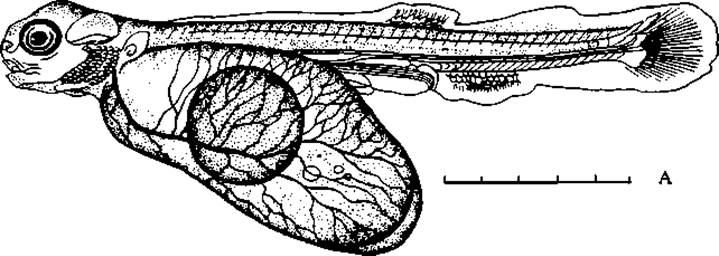
Рис. 58. Свободный эмбрион горбуши с желточным мешком. А — абсолютная величина эмбриона
Кроме искусственного рыборазведения описанным способом р низовьях рек (например, Волги) улучшают условия нереста полупроходных рыб (например, сазана, леща). В таких нерестововыростных хозяйствах используют естественные водоемы или создают искусственные путем обваловывания близкого к реке участка суши площадью в несколько сот гектаров. Весной такой участок заполняют водой, поступающей из реки по специальному каналу. Водоемы зарыбляют отловленными в реке производителями из расчета на 1 га 8 экземпляров сазана и 30—40 экземпляров леща при соотношении полов 1 : 1. На этих нерестилищах происходят естественное размножение рыб и рост их мальков. В конце лета открывают нижний шлюз и мальки вместе со спускаемой водой выходят в реку. С 1 га сбрасывают в реку примерно 50 тыс. мальков сазана (массой 3 г) и 80 тыс. мальков леща (массой 0,5 г).
При сокращении мест естественных нерестилищ создаются разного типа искусственные нерестилища. Для рыб, откладывающих икру на растения (фитофильные рыбы), искусственные нерестилища могут быть стационарными или плавучими. Последние имеют особое значение в условиях сильного колебания уровня воды, например на водохранилищах при гидростроительстве.
Принимаются меры и к улучшению условий на естественных нерестилищах. Так, например, в ряде районов Дальнего Востока в суровые малоснежные зимы происходят массовое промерзание нерестовых бугров (гнезд) и гибель находящейся в них икры лососей. Борьба с этим явлением возможна путем создания более высокого уровня воды в реке зимой и строительства после окончания нереста временных подпорных плотин, снегозадержания на льду нерестового водоема и ряда других мер.
Основным объектом прудового рыборазведения служит карп — одомашненная форма сазана. Карпов разводят в системе прудов, имеющих разное назначение. Нерестовые пруды небольшие — площадью около 1 000 м2, глубиной 0,5— 1,2 м, со свежей зеленой Растительностью, на которую карпы откладывают икру. Когда молодь Несколько подрастет (через 10—15 дней после выхода из икры), ее переводят в большие и более глубокие выростные пруды из расчета 12 тыс. штук на 1 га пруда. Перезимовавших годовиков держат в нагульных прудах (400 — 500 штук на 1 га). Зимние пруды, или зимовальники, служат для зимнего содержания производителей и сеголеток. Эти пруды делают наиболее глубокими (более 3 м), чтобы они не промерзали. Вода в них должна быть проточной и полностью обновляться в течение 5 — 8 суток. В зимовальнике карпы не питаются.
Кроме комплексных хозяйств, карпа широко разводят у нас в так называемых однолетних нагульных хозяйствах. При этом молодых рыб, выращенных в специальных питомниках, выпускают в нагульные водоемы, где они растут до осени, после чего пруды спускают или рыб вылавливают неводом. При правильной постановке дела прудовое карповое хозяйство может дать на 1 га водного зеркала нагульного пруда до 600 трехлеток массой 1 — 1,5 кг.
Местами практикуется совместное разведение карпа и растительноядных рыб (белого амура, толстолобика). Есть удачные опыты совместного разведения карпа и щук, которые существуют за счет избыточной молоди карпа и тем самым повышают общую рыбопродуктивность водоема.
Акклиматизация. Значительное развитие получили в России работы по искусственному обогащению промысловой фауны рыб путем акклиматизации многих ценных видов.
Так, в 30-х гг. XX столетия из Черного моря в Каспийское было пересажено около 3 млн особей кефали. Рыба эта успешно акклиматизировалась.
Сиги, обитающие в озерах северо-западных областей, успешно акклиматизированы в озерах Урала, Зауралья, Красноярского края. Местами в результате этого мероприятия рыбная продуктивность водоемов увеличилась в несколько раз.
В одном из наиболее крупных озер Средней Азии — Иссык-Куле — успешно акклиматизирована форель, вывезенная из озера Севан. Характерно, что в водоемах бассейна Иссык-Куля форель растет быстрее и достигает больших размеров. Там же хорошо акклиматизировался лещ.
Большие и успешные работы проведены по искусственному расселению сазана. Он акклиматизирован в водоемах Ленинградской, Новгородской областей, в озерах Зауралья и в других местах.
Удачно акклиматизированы тихоокеанские лососи у Атлантического побережья Северной Америки и Скандинавии. Предприняты удачные попытки акклиматизации растительноядных рыб Юго-Восточной Азии — белого амура и толстолобика. Эти мероприятия преследовали одновременно две цели: повышение рыбопродуктивности водоемов и борьбу с их зарастанием. Белый амур акклиматизирован также на Волге, Кубани, в Цимлянском и других водохранилищах.
Промысловый эффект акклиматизации показывает долевое значение акклиматизированных рыб в улове: в Курганской области оно составило 4 %, в Свердловской — 10 %, в Челябинской — 20 % от всего улова.
Акклиматизируют и разводят рыб и с целью борьбы с комарами. Мелкая американская рыбка гамбузия была завезена в пределы России в 1924 г. Она широко распространилась по южным озерам и рекам, активно питаясь личинками комаров.
В 1970-х гг. в Сибирь из Северной Америки ввезены для акклиматизации 3 вида буффало (Ictiobus) — крупных рыб из отряда Карпообразные с большим выходом товарной массы и т.д.
Филогения низших черепных
Численность многих популяций рыб стала чрезвычайно низкой из-за перепромысла, строительства гидросооружений, загрязнения водоемов. Так, в Красную книгу РФ включен 21 вид и подвид рыб. Особой охраны и специальных мер по восстановлению природных популяций требуют 11 видов осетрообразных, особенно белуги (Huso huso) и калуги (Huso dauricus). Из-за перепромысла подорваны популяции многих лососеобразных (Salmoniformes), из них 11 видов и подвидов требуют специальных мер охраны.
Среди современных черепных (или позвоночных) животных наиболее примитивны Круглоротые (Cyclostomata). Однако хорошо известно, что они являются в то же время и весьма специализированными. К сожалению, палеонтология не дает прямого ответа на вопрос об особенностях организации предков позвоночных.
Сохранение остатков ранних позвоночных в ископаемом состоянии в палеонтологической летописи приурочено к тому историческому периоду, когда у них возник костный скелет. Самых древних примитивных позвоночных объединяют в надкласе Бесчелюстные (Agnatha) под названием Ostracodermii («Раковинокожие», или Щитковые).
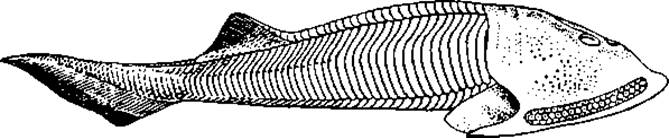
Рис. 59. Щитковое (Cephalaspis Lyelli) из нижнего девона
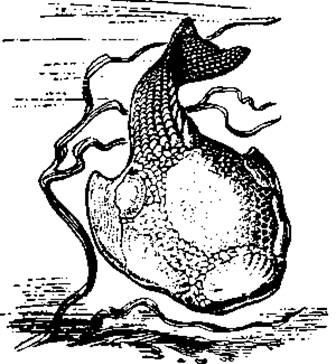
Рис. 60. Щитковое (Psammolepis venlukovi) из девонского периода
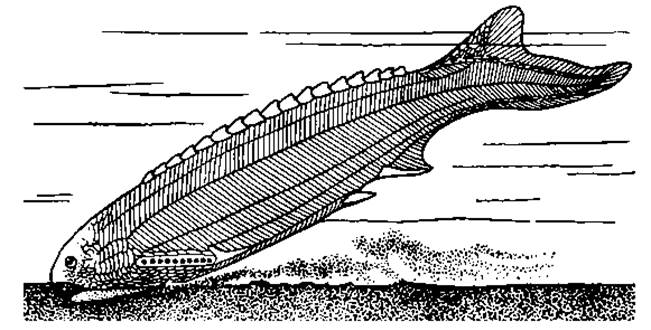
Рис. 61. Щитковое и шрафт (Anaspida, Pterolepis hitida) силурского периода
Они обладали мощным наружным скелетом в виде или сплошного панциря, или отдельных крупных костных пластин, мелких чешуй, или, наконец, сочетания всех этих образований. Некоторые остракодермы с тяжелым скелетом, дорсовентрально уплощенные, часто с гипоцеркальным хвостовым плавником, вели, видимо, придонный образ жизни. Другие — с гибким веретеновидным телом, покрытым мелкими чешуями, вероятно, обитали в толще воды (рис. 59, 60, 61). Щитковые в целом были разнообразной группой, включавшей так называемых костнощитковых, разнощитковых, бесщитковых и прочих. Первые описанные формы были обнаружены в отложениях ордовикского периода. Своего расцвета они достигли в силуре — девоне. К концу девона большая часть их вымерла, предположительно из-за конкуренции со стороны челюстноротых, разнообразие которых к этому времени возросло. В последующей палеонтологической летописи их останки не обнаружены. Вместе с тем ряд черт сближает эту группу с современными круглоротыми — миногами и миксинами. Предполагают, что обе группы современных бесчелюстных могли появиться в карбоне. Со щитковыми их сближает сходство организации: отсутствие челюстей и парных конечностей, рот сосущего типа, два полукружных канала во внутреннем ухе, внутренний скелетный тяж — хорда, энтодермальные жабры, непарный обонятельный орган. Бесчелюстные составили одну весьма примитивную и специализированную эволюционную линию позвоночных.
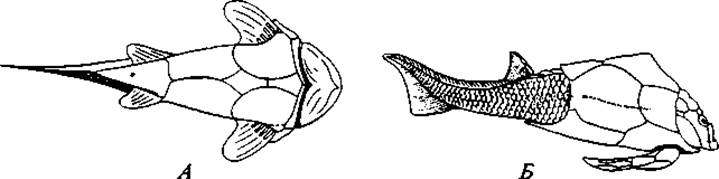
Рис. 62. Панцирные рыбы:
А — Coccosteus из среднего девона (вид снизу); Б — Pterichthyes
Рис. 63. Cladoselache — верхнедевонская акулоподобная рыба
Другая, ведущая, ветвь представлена Челюстноротыми (Gnathostomata — Ectobranchiata). Она дала рыб и всех более высокоорганизованных позвоночных. Непосредственные предки рыб пока неизвестны. Самые ранние ископаемые останки их в виде чешуй обнаружены в верхнесилурийских отложениях. В девонских отложениях находят уже представителей весьма разнообразных групп. Одной из наиболее ранних групп, известных с раннего силура, являются Панцирные рыбы (Placodermi), их тело было покрыто костным панцирем (рис. 62). Наряду с примитивными чертами они имели некоторые прогрессивные особенности: внутренний скелет их состоял главным образом из хряща, и они имели костные челюсти. Парные плавники (чаще только передние) были расчленены на отделы и прикрыты костными пластинками. Панцирные рыбы обитали как в пресных водах, так и в солоноватых. Они дожили до каменноугольного периода и затем вымерли. Другой группой, столь же древней (силур) и примитивной, были мелкие пресноводные акантодии (Acanthodii), тело которых было покрыто ганоидной чешуей. Плавники их имели широкое основание, и между грудными и брюшными плавниками находились ряды добавочных маленьких плавников, также с широкими основаниями. В этом можно видеть остатки некогда сплошных складок, находившихся на боках тела и давших начало парным конечностям. Кроме того, на примитивность описываемых рыб указывает членистость их челюстных дуг. т. е. большее, чем у других рыб, сходство челюстных дуг с другими висцеральными дугами.

Рис. 64. Palaeoniscus из пермских отложений (длина тела до 25 см)
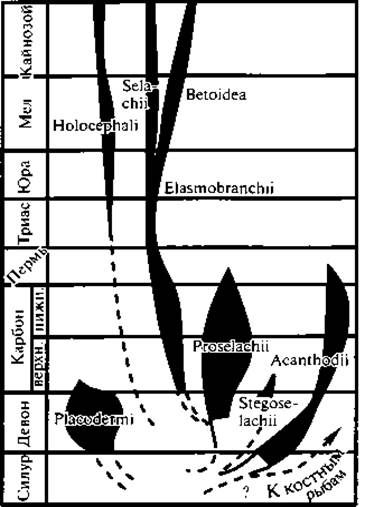
Рис. 65. Филогенетическое древо панцирных и хрящевых рыб
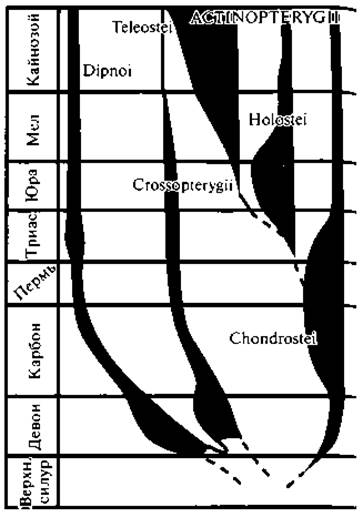
Рис. 66. Филогенетическое древо костных рыб
Упомянем о подклассе Первичные акулы (Proselachii), из которых хорошо известна позднедевонская Cladoselache (рис. 63).
Собственно Хрящевые рыбы (Chondrichthyes) известны с раннего девона. Пластинчатожаберные (Elasmobranchii) пережили два всплеска адаптивной радиации — в силуре — девоне и в раннем мезозое. С конца мезозоя оформились современные семейства этого подкласса. Цельноголовые (Holocephalia), известные с раннего карбона, никогда не были многочисленными.
Класс Костные рыбы (Osteichthyes) рано оформился в две группы: подкласс Лопастепёрые (Sarcopterygii) и подкласс Лучепёрые (Actinopterygii).
Наиболее древними лучепёрыми были Палеонисциды (Palaeoniscoidei) (рис. 64). Они характеризуются наличием главным образом наружных кожных окостенений, гетероцеркальным хвостом, наличием рострума, ганоидной чешуей и присутствием V-образных чешуек — фулькры на спинном и хвостовом плавниках. Все это дает повод считать их весьма близкими к современным хрящекостным, в частности к осетровым рыбам. От палеонисцид ведут свое начало и другие ганоидные (Ganoidomorha), возникшие в триасе и занимающие промежуточное положение между палеонисцидами и костистыми рыбами. В середине мезозоя они были господствующей группой рыб, но к настоящему времени сохранились лишь два представителя — амия и кайманова рыба.

Рис. 67. Кистепёрая рыба (Undina pencillata) из верхней юры
Собственно Костистые рыбы (Teleostei) возникли в мезозое. Их эволюция шла быстро и многообразно. В настоящее время это господствующая группа рыбы (рис. 65, 66).
Подкласс Лопастепёрые (Sarcopterygii) содержит надотряды Кистепёрые (Crossopterygii) и Двоякодышащие (Dipnoi) рыбы. Предки кистепёрых и двоякодышащих, несомненно, были близки между собой. Обе группы появляются в нижнем девоне и расцвета достигают в верхнем девоне и каменноугольном периоде. Дифференцировка этих двух групп легочнодышащих рыб была связана в значительной мере с характером питания. В то время как кистепёрые (рис. 67) оставались хищниками и сохранили способность быстро плавать и хватать добычу, двоякодышащие перешли к питанию придонными беспозвоночными, они утратили хорошо развитые плавники и стали малоподвижными, что мы и наблюдаем у немногих современных представителей.
Кистепёрые рыбы представляют собой интерес в связи с тем, что из всех рыб они ближе всего стоят к исходной группе наземных позвоночных — Панцирным амфибиям (Stegocephalia).
Рыбы возникли в пресных водоемах и только в последующем, хотя и довольно быстро, заселили Мировой океан.