Химия белка - Часть 1 - Общая химия белка - Ашмарин И. П 1968
Пространственная организация белковой молекулы
Методика изучения вторичной структуры белков и полипептидов
Вращение плоскости поляризации света (оптическая активность)
Из опыта хорошо известно, что все белки и синтетические полипептиды вращают плоскость поляризации света, т. е. оптически активны. Наблюдаемая оптическая активность имеет составной характер. Это прежде всего сумма вкладов, вносимых асимметрическими а-углеродными атомами аминокислот. Поскольку все природные аминокислоты левовращающие, то и в полипептидной цепи а-С-атомы вращают влево. Следовательно, знак этого слагаемого общей оптической активности отрицателен, и любой полипептид, находящийся в форме беспорядочного клубка, будет обладать отрицательной активностью.
Однако в случае белков или полипептидов с большой степенью спирализации возникает второй большой вклад в оптическое вращение — за счет внутренней асимметрии самой а-спирали. Как уже говорились, полипептиды, построенные из L-аминокислот, образуют, вероятно, только правые спирали, которые будут вращать плоскость поляризации вправо. Таким образом, второй инкремент оптической активности по знаку противоположен первому; по абсолютным величинам эти компоненты довольно близки друг к другу, и в целом величина оптической активности полипептидной спирали приближается к нулю. Каким же образом можно оценить эти составные оптические активности и на основании получаемых данных рассчитать степень спирализации полипептидной цепи?
Вращательную способность какого-либо соединения в виде чистого твердого вещества, жидкости или раствора выражают через удельное вращение [а]D (удельная активность, измеряемая для D-линии натрия, 589,3 ммк), определяемое по уравнению
![]()
где a — измеренный угол вращения; I — толщина слоя в дм; с — концентрация оптически активного вещества в г/100 мл.
Имея дело с полипептидами и другими соединениями с длинной цепью, удобнее выражать оптическую активность вращением звена цепи («вращением остатка»):
![]()
где М0 — молекулярный вес звена, т. е. аминокислотного остатка.
Сравнивая вращение полипептидов в различных растворителях, важно учесть неспецифический эффект показателя преломления n. Зависимость удельного вращения от показателя преломления имеется даже тогда, когда структура вещества нечувствительна к растворителю. Поэтому целесообразнее приводить величину удельного вращения или вращения остатка к условной среде с n = 1. В этом случае мы будем иметь скорректированное вращение остатка, или «эффективное вращение звена цепи»:
![]()
Величину [m] для полипептида в форме спирали или клубка можно легко рассчитать, если нам известны молекулярный вес аминокислотного остатка и удельное вращение [ah для спиральной и клубковой форм. Последнее определяют в опытах по плавлению а-спиралей синтетических полипептидов. Так, если оттитровывать боковые карбоксильные группы полиглютаминовой кислоты щелочью, то нейтральные СООН-группы будут переходить в заряженные СОО--группы. В результате вдоль молекулы начнут действовать мощные силы электростатического отталкивания, способные разорвать водородные связи и полностью расплавить а-спираль (рис. 2І). Это разрушение а-спирали происходит в узкой зоне pH и сопровождается резким изменением оптической активности. Удельное вращение теряет большой положительный инкремент, создаваемый а-спиралью, и падает от +6 до —80°. Так как спираль полностью разрушена, то эта величина, —80°, и есть удельное вращение клубковой формы, обусловленное а-углеродными атомами L-аминокислот. Соответственно вклад а-спирали гораздо больше и обратен по знаку, составляя +86°. Алгебраическая же сумма обеих величин, т. е. [a]D спиральной конфигурации, близка к нулю (+ 6°).
Довольно близкие значения были получены и для величины [m]. Так, для синтетических полипептидов в форме клубка и для денатурированных белков эффективное вращение среднего аминокислотного звена колебалось от —80 до —110°, тогда как у полностью спирализованных полипептидов и нативных белков эта величина была близка к нулю. Следовательно, положительный инкремент величины [а]D или [m], относящийся к идеальной a-структуре, составляет 80—100°. По данным для синтетических полипептидов и парамиозина, чья структура считается полностью спирализованной, этот вклад равен 90°. Таким образом, помещая полипептид в различные растворители (например, от чистого дихлорэтана до его смеси с дихлоруксусной кислотой и самой кислоты), из изменения величины [а]D или [m] мы можем приблизительно оценить процент а-спиральности цепи.
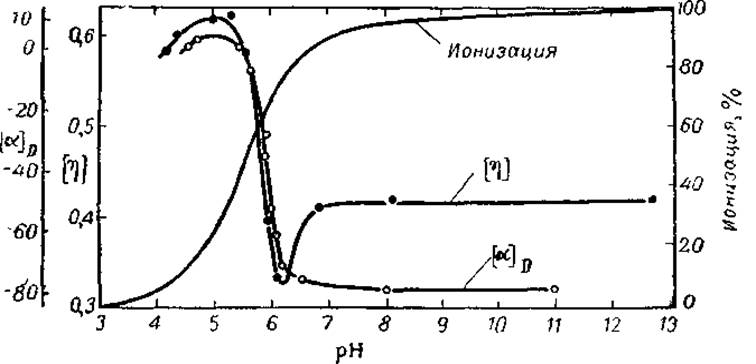
Рис. 21. Плавление спиралей полиглютаминовой кислоты при изменении степени ионизации карбоксильных групп (по Доти, 1961).
Значит ли это, что таким критерием, как оптическая активность, можно уверенно пользоваться не только для суждения о наличии спиральных участков в белковой молекуле, но и для оценки степени спирализации? К сожалению, нет. Точное доказательство наличия а-спирали может быть получено лишь на основании рентгеноструктурного анализа, тогда как большой положительный вклад в оптическую активность можно с уверенностью приписать лишь наличию в полипептиде или глобулярном белке какой-то упорядоченной структуры. Так, при помещении полипептида поли-О-ацетилсерина из хлороформа в дихлоруксусную кислоту также происходит переход к более отрицательным величинам [a]D. Однако методом инфракрасной спектрофотометрии было показано, что этот полипептид дает только ß-структуры. Следовательно, упорядоченной структурой, обусловливающей положительный инкремент [a]D, может являться не только а-спираль, но и ß-структура. Оценка степени спирализации по величине оптической активности также весьма условна. Так, при плавлении вторичной структуры трипсина и одного из его фрагментов величины [а]D изменяются соответственно с —35 до —55° и с —75 до —40°. Это говорит о том, что фрагмент трипсина состоит преимущественно из левых спиралей, тогда как в макромолекуле трипсина преобладают в целом правовращающие структуры. Отсюда ясно, что спиральные участки в трипсине могут давать слагаемые разного знака, и величина оптической активности оказывается по абсолютной величине меньшей, чем в случае белка с той же степенью спирализации, но с одними правыми спиралями. Аналогичные правые (цепь В) и левые (цепь А) спирали были обнаружены и в молекуле инсулина.
Таким образом, само по себе изменение оптической активности, сопровождающее распад упорядоченной структуры, не есть доказательство существования спиралей в составе полипептида или белка. Оно должно быть дополнено данными па скорости обмена имидного водорода, поглощению в ультрафиолетовом свете и другими.