Биохимические основы жизнедеятельности организма человека - Волков Н.И., Несен Э.Н. 2000
Биохимия спорта
Биохимические изменения в организме при выполнении упражнений различной мощности и продолжительности
Общая направленность изменения биохимических процессов при мышечной деятельности
Степень изменения биохимических процессов в организме при мышечной деятельности зависит от типа выполняемого упражнения, его мощности и продолжительности, а также от тренированности спортсмена. В первую очередь такие изменения касаются механизмов аэробного и анаэробного энергообразования, что наиболее наглядно прослеживается на примере энергообеспечения бега на разные дистанции, представляющего собой работу различной мощности и продолжительности (предельное время представлено на рис. 131).
При беге на 100 м энергетическое обеспечение работы мышц осуществляется только за счет анаэробных процессов (рис. 133). На первых метрах дистанции или стартовом разбеге преимущественное значение в энергообеспечении работающих мышц имеет креатинфосфокиназный механизм. На остальной части дистанции поддержание достигнутой максимальной скорости бега обеспечивается одновременным использованием креатинфосфата и гликогена.
Общая доля анаэробного гликолиза в энергетике спринтерского бега составляет около 50 %. Скорость гликолитического распада гликогена при беге с максимальной скоростью на дистанции 100 м может достигать 1000-кратного увеличения по сравнению с уровнем покоя. Однако запасы гликогена не исчерпываются даже при интенсивной работе до изнеможения, что следует из табл. 25, в которой представлены значения концентрации АТФ, КрФ и гликогена в мышце человека, определенные на материале, полученном путем биопсии, в состоянии покоя и после максимальной нагрузки различной продолжительности. Содержание АТФ изменяется незначительно, а концентрация КрФ снижается от 17 ммоль ⋅ кг-1 влажной массы мышцы почти до нуля после максимальной нагрузки до изнеможения.
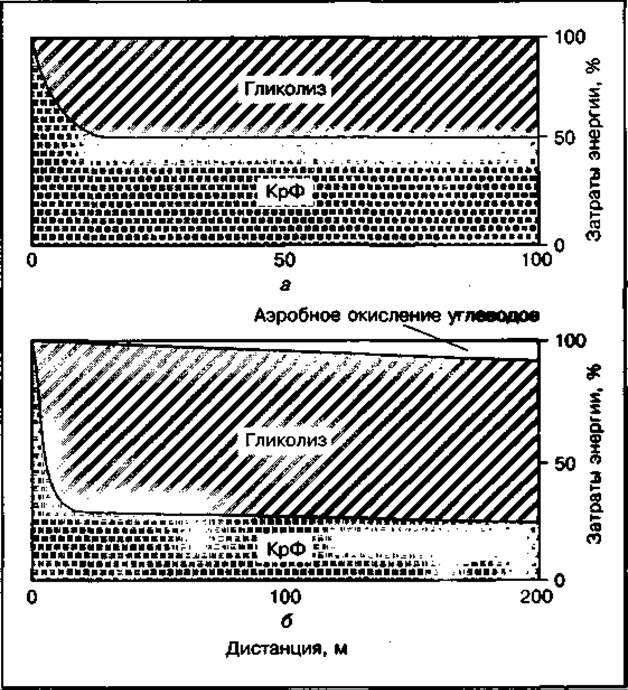
Рис. 133 Механизмы энергообеспечения бега на 100 м (а) и 200 м (б)
ТАБЛИЦА 25 Концентрация энергетических субстратов и pH в скелетных мышцах при выполнении физической работы
Показатели |
В состоянии покоя |
После 6—10 с физической работы |
После 30 с физической работы |
В состоянии изнеможения |
АТФ |
5 |
5 |
3 |
3 |
КрФ |
17 |
12 |
5 |
1 |
Гликоген |
85 |
74 |
68 |
50 |
Лактат |
2 |
7 |
22 |
32 |
pH |
7,0 |
6,9 |
6,7 |
6,3 |
Примечание. Представлены усредненные данные многих исследователей. Метаболиты мышцы измерялись в миллимолях на килограмм влажной массы тела
При беге на 200 м креатинфосфат и анаэробный гликолиз также используются одновременно при заметном преобладании гликолиза в общей энергетике работы. Однако уже на 150-м метре запасы креатинфосфата в работающих мышцах заметно истощаются и темп бега обычно снижается примерно на 10 %, при этом в энергетическом обеспечении бега определенную роль начинает играть аэробный метаболизм. Общий вклад аэробного способа производства энергии на этой дистанции может составить 10-20 %.
При беге на 400 м все три источника энергии участвуют в поддержании заданной скорости бега. Примерно 10% общих затрат энергии обеспечивает реакция распада креатинфосфата. Она играет основную роль в период стартового разгона и на финише бега. Наибольшее значение в энергетике бега на 400 м имеет анаэробный гликолиз, обеспечивающий свыше 60 % энергии, затрачиваемой на преодоление дистанции. На долю аэробного процесса приходится около 25 % общего энергетического запроса (рис. 134).
Основными факторами, ограничивающими скорость бега на дистанции 400 м и способствующими развитию утомления, является ограниченная емкость креатинфосфокиназного механизма, а также значительное накопление молочной кислоты в работающих мышцах и закисление внутренней среды организма. Увеличение в процессе тренировки доли аэробного ресинтеза АТФ за счет окисления углеводов может рассматриваться как перспективный путь повышения выносливости бегунов, специализирующихся на этой дистанции.
При беге на средние дистанции (800 и 1500 м) креатинфосфокиназный механизм и анаэробный гликолиз вносят определенный вклад в ресинтез АТФ при работе, однако аэробный метаболизм становится доминирующим. На его долю приходится около 45 % общих затрат энергии на дистанции 800 м и до 75 % — на дистанции 1500 м (рис. 135). Важное значение при этом имеют запасы гликогена в работающих мышцах, которых может хватить при анаэробном окислении примерно на 1000 м. Для повышения выносливости в данных видах бега более эффективно использовать эти запасы гликогена в ходе аэробных окислительных процессов. Это потребует усиленного кровоснабжения работающих мышц и потребления кислорода мышцами, а также увеличения буферных резервов крови, что будет предотвращать значительное закисление и раннее развитие утомления.
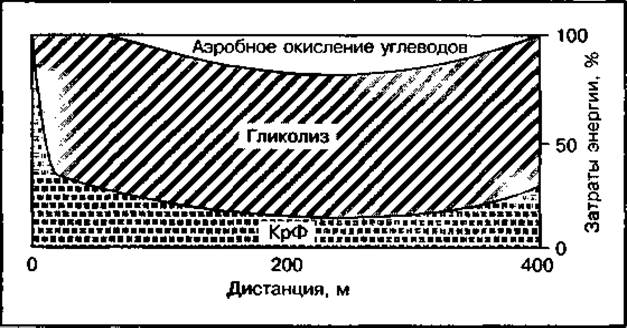
Рис. 134 Механизмы энергообеспечения бега на 400 м
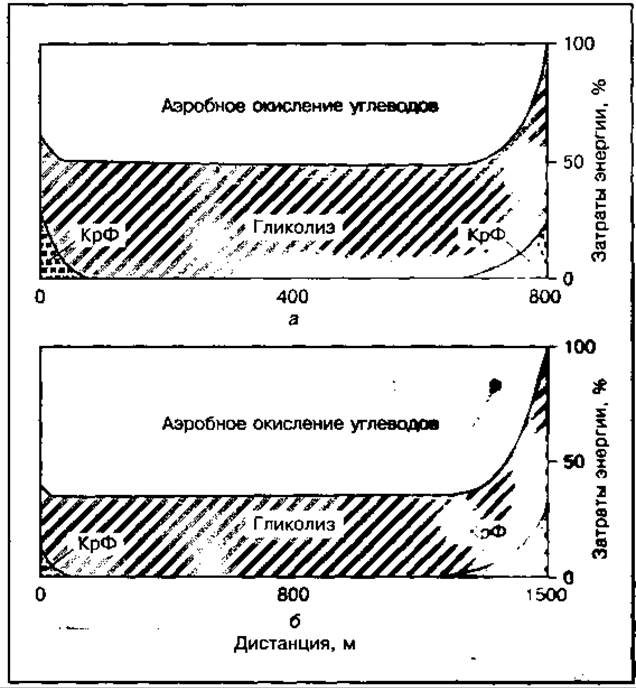
Рис. 135 Механизмы энергообеспечения бега на 800 м (а) и 1500 м (б)
При беге на длинные дистанции (5 и 10 км) аэробное окисление углеводов является основным механизмом энергообеспечения работы, так как на его долю приходится до 87 % общих затрат энергии на дистанции 5 км и около 97 % — на дистанции 10 км (рис. 136). На этих дистанциях вклад анаэробных источников энергии также еще достаточно большой. Он может достигать 15 % общих затрат энергии и играет важную роль при финишном ускорении, приносящем победу при беге на длинные дистанции. Наиболее значительным фактором, влияющим на работу на выносливость, является кислородное снабжение работающих мышц, поскольку потребление кислорода во время бега поддерживает максимальную скорость окисления углеводов. Порог анаэробного обмена у стайеров при работе 75—90 % МПК.
При марафонском беге затраты энергии восполняются исключительно за счет аэробного процесса. Погашение этих затрат невозможно только за счет окисления углеводов из-за недостаточности запасов гликогена в работающих мышцах спортсмена, поэтому значительная часть энергии образуется за счет окисления жиров, на долю которых может приходиться от 10 до 50 % общих затрат энергии (рис. 137). Вклад жиров в энергообеспечение при мышечной работе на длинных и сверхдлинных дистанциях у высокотренированных бегунов с большими запасами гликогена в работающих мышцах составляет 12—20%, у нетренированных бегунов — более 60 %. Всего на дистанции марафонского бега окисляется около 300 г жиров. Использование жиров в качестве источника энергии менее эффективно сравнению с окислением углеводов, так как происходит оно с более низкой скоростью и с большим потреблением кислорода. Увеличивающаяся концентрация жирных кислот в крови при мобилизации жиров и высокая скорость их окисления в работающих мышцах тормозят окисление в них углеводов, в основном глюкозы крови, что влечет за собой снижение скорости бега. Увеличение доли окисляемых жиров при работе способствует также развитию центрального торможения. Мобилизация жиров при физической работе активизируется рядом гормонов (рис. 138).
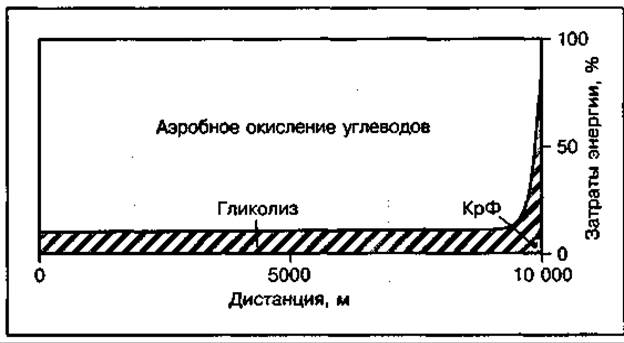
Рис. 136 Механизмы энергообеспечения бега на 10 000 м
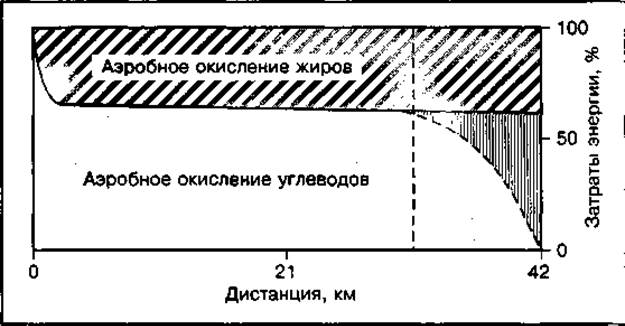
Рис. 137 Механизмы энергообеспечения марафонского бега (указан момент, когда вследствие исчерпания запасов гликогена резко усилилось окисление жиров в работающих мышцах)
При длительной работе наряду с увеличением использования в энергетическом обмене жиров может происходить новообразование углеводов из веществ неуглеводной природы (глюконеогенез), активируемое гормонов кортизолом. Основным субстратом глюконеогенеза являются аминокислоты часть которых накапливается в мышцах при работе в результате распад тканевых белков. Они могут быть использованы для образования глюкозы.
Данные об участии различных источников энергии в обеспечении ресинтеза АТФ при беге на разные дистанции приведены в табл. 26. Вклад отдельных энергетических источников в обеспечение работы имеет важное значение при выборе оптимальной стратегии подготовки в избранном виде спорта, в том числе близких по интенсивности и продолжительность к рассмотренным физическим беговым нагрузкам.
Представление об использовании отдельных механизмов энергообеспечения при мышечной работе можно получить по результатам изменения концентрации молочной кислоты и глюкозы в крови. Как следует из рис. 139, максимальное накопление молочной кислоты в крови достигается при беге на 400 и 800 м, что свидетельствует об использовании лактатного механизма ресинтеза АТФ, а глюкозы — при беге на 10 000 м, когда аэробный механизм ресинтеза АТФ становится ведущим.
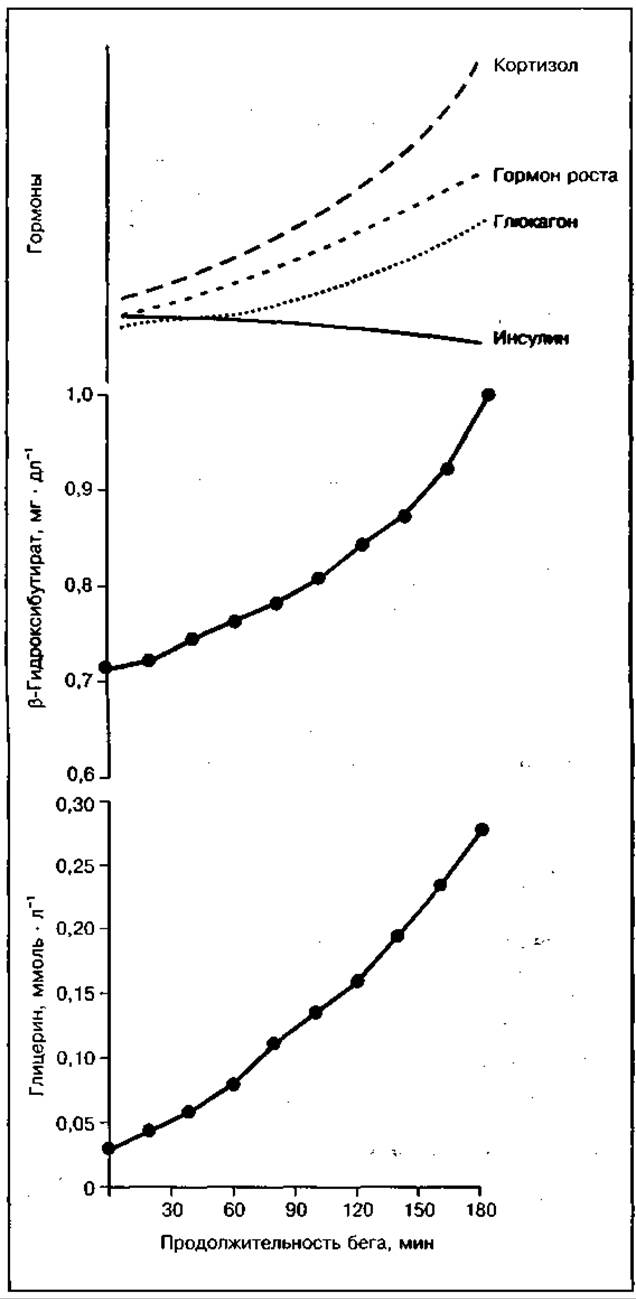
Рис. 138 Изменение концентрации продуктов распада жиров (глицерина, ß-гидроксибутирата) и гормонов, активирующих липолиз, при выполнении упражнений разной продолжительности
ТАБЛИЦА 26. Участие различных источников энергии в обеспечении ресинтеза АТФ при беге на различные дистанции
|
Вклад в генерирование АТФ различных механизмов ее ресинтеза, % |
|||||
Дистанция |
Креатинфосфат |
Окисление гликогена мышц |
Глюкоза крови (гликоген печени) |
Триглицериды (жирные кислоты) |
|
анаэробное |
аэробное |
||||
100 м |
50,0 |
50,0 |
— |
— |
— |
200 м |
25,0 |
65,0 |
10,0 |
— |
— |
400 м |
12,5 |
62,5 |
25,0 |
— |
— |
800 м |
Минимальный |
50,0 |
50,0 |
— |
— |
1 500 м |
" |
25,0 |
75,0 |
— |
— |
5 000 м |
" |
12,5 |
87,5 |
— |
— |
10 000 м |
" |
3,0 |
97,0 |
— |
— |
Марафон |
— |
— |
75,0 |
5 |
20 |
Супермарафон (84 км) |
— |
— |
35,0 |
5 |
60 |
24-часовый забег |
— |
— |
10,0 |
2 |
88 |
Примечание. На дистанциях 800, 1500, 5000 и 10 000 м креатинфосфат используется только на первых секундах бега или на финишном отрезке дистанции
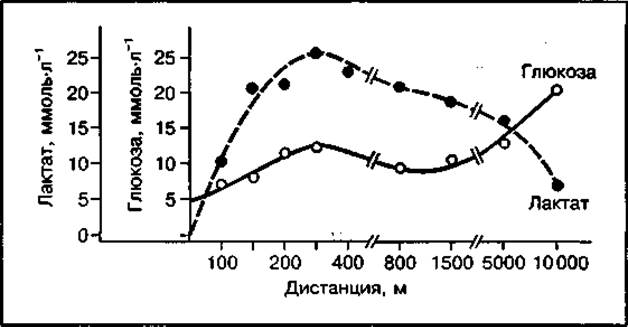
Рис. 139 Изменение концентрации лактата и глюкозы в крови спортсменов после бега на дистанции от 100 до 10 000 м