Биологическая химия - Березов Т. Т., Коровкин Б. Ф. 1998
Метаболизм углеводов
Пентозофосфатный путь окиснения углеводов
Регуляция метаболизма углеводов
Пути регуляции метаболизма углеводов крайне разнообразны. На любых уровнях организации живого организма обмен углеводов регулируется факторами, влияющими на активность ферментов, участвующих в реакциях углеводного обмена. К этим факторам относятся концентрация субстратов, содержание продуктов (метаболитов) отдельных реакций, кислородный режим, температура, проницаемость биологических мембран, концентрация коферментов, необходимых для отдельных реакций, и т.д. (см. главу 4).
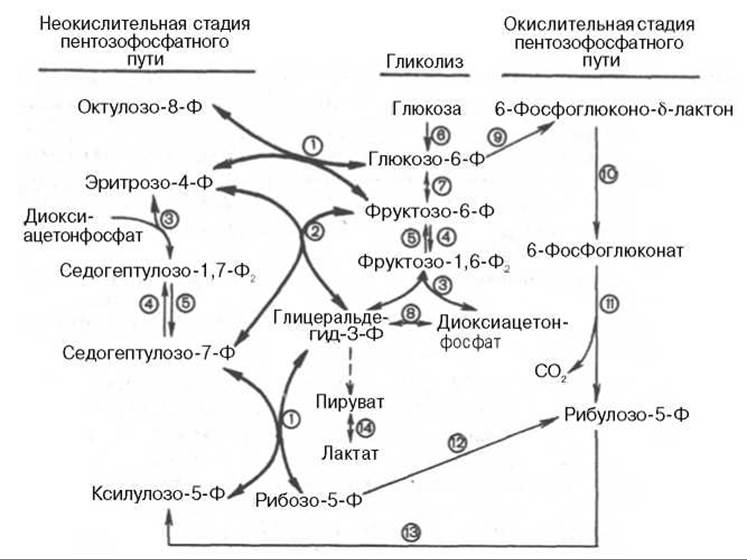
Рис. 10.13. Современная схема пентозофосфатного пути окисления углеводов, отражающая его связь с гликолизом (по Херсу).
1 - транскетолаза; 2 - трансальдолаза; 3 - альдолаза; 4 - фосфофруктокиназа; 5 - фруктозо-1,6-бисфосфатаза; 6 - гексокиназа; 7 - глюкозофосфатизомераза; 8 - триозофосфатизомераза; 9 - глюкозо-6-фосфатдегидрогеназа; 10 - 6-фосфоглюконолактоназа; 11 - 6-фосфоглюконатдегидрогеназа; 12 - изомераза; 13 - эпимераза; 14 - лактатдегидрогеназа.
В данной главе было показано влияние перечисленных факторов на активность ферментных систем углеводного обмена. И тем не менее некоторые аспекты регуляции метаболизма углеводов напомним.
Гликолиз — это совокупность реакций превращения глюкозы в пируват. У аэробных организмов гликолиз служит как бы прелюдией к циклу трикарбоновых кислот (циклу Кребса). Десять реакций гликолиза протекают в цитозоле. Гликолитический путь играет двоякую роль: приводит к генерированию АТФ в результате распада глюкозы, и он же поставляет строительные блоки для синтеза клеточных компонентов. Реакции гликолитического пути в физиологических условиях легкообратимы, кроме реакций, катализируемых гексокиназой, фосфофруктокиназой и пируваткиназой. Фосфофруктокиназа — наиболее важный регуляторный элемент (фермент) в процессе гликолиза, ингибируется высокими концентрациями АТФ и цитрата и активируется АМФ.
Цикл трикарбоновых кислот (цикл Кребса) представляет собой конечный общий путь для окисления «топливных» молекул. Большинство «топливных» молекул вступает в цикл в виде ацетил-КоА. Окислительное декарбоксилирование пирувата, приводящее к образованию ацетил-КоА, является связующим звеном между гликолизом и циклом трикарбоновых кислот. Заметим, что последний служит также источником строительных блоков для процессов биосинтеза. Все реакции цикла протекают в митохондриях.
Скорость цикла трикарбоновых кислот зависит от потребности в АТФ. Высокий энергетический заряд клетки понижает активность цитратсинтазы, изоцитратдегидрогеназы и а-кетоглутаратдегидрогеназы. Еще один важный регуляторный момент — необратимое образование ацетил-КоА из пирувата. В результате пентозофосфатного пути происходит генерирование НАДФН и рибозо-5-фосфата в цитозоле. НАДФН участвует в восстановительных биосинтезах, а рибозо-5-фосфат используется в синтезах РНК, ДНК и нуклеотидных коферментов.
Взаимодействие гликолитического и пентозофосфатного путей обеспечивает возможность постоянного приспособления концентраций НАДФН, АТФ и строительных блоков, например рибозо-5-фосфата и пирувата, для удовлетворения потребностей клеток.
Наконец, глюконеогенез и гликолиз регулируются реципрокно, так что, если активность одного из путей относительно понижается, то активность другого пути повышается.
У человека и животных на всех стадиях синтеза и распада углеводов регуляция углеводного обмена осуществляется при участии ЦНС и гормонов.
Например, установлено, что концентрация глюкозы в крови ниже 3,3—3,4 ммоль/л (60—70 мг/100 мл) приводит к рефлекторному возбуждению высших метаболических центров, расположенных в гипоталамусе. В регуляции углеводного обмена особая роль принадлежит высшему отделу ЦНС — коре большого мозга. Наряду с ЦНС важное влияние на содержание глюкозы оказывают гормональные факторы, т.е. регуляции уровня глюкозы в крови осуществляется ЦНС через ряд эндокринных желез (см. главу 8).