Биологическая химия - Березов Т. Т., Коровкин Б. Ф. 1998
Обмен простых белков
Переваривание белков
Эндопептидазы
Пепсин. Одним из хорошо изученных и основных протеолитических ферментов пищеварительного тракта является пепсин. Его наличие в желудке было установлено еще в 1783 г. Л. Спалланцани, хотя в кристаллическом виде он был получен только в 1930 г. (см. главу 1). Пепсин вырабатывается в главных клетках слизистой оболочки желудка в неактивной форме — в виде пепсиногена. Превращение пепсиногена в активный пепсин происходит в желудочном содержимом, однако молекулярный механизм этого превращения в деталях еще не выяснен. Наиболее вероятным считается предположение, что этот процесс является последовательным и протекает в несколько этапов в присутствии соляной кислоты по механизму аутокаталитического действия самого пепсина. Молекулярная масса пепсиногена составляет приблизительно 40400, а пепсина — 32700, поэтому превращение первого во второй связано с отщеплением пептидных фрагментов. Оба фермента можно сравнительно легко получить в кристаллическом виде. Следует отметить, что в отличие от других протеиназ пепсин отличается высокой устойчивостью в сильнокислой среде и характеризуется низким значением изоэлектрической точки (рI < 1). Такие условия обычно создаются в желудочном содержимом, куда поступает секретируемая париетальными клетками слизистой оболочки соляная кислота*; рН чистого желудочного сока колеблется от 1,0 до 2,0. Эта среда является оптимальной для каталитического действия пепсина. Имеются доказательства, что в желудке человека из пепсиногена, вероятно, образуется не только активный пепсин, а несколько близких по строению пепсинов, включая пепсиноподобный фермент гастриксин, который имеет отличный от пепсина оптимум рН действия, равный 3,0.
Реннин. Фермент реннин выделен из сока четвертого отдела желудка телят в кристаллическом виде. Он есть также в желудочном соке детей грудного возраста. По механизму и специфичности действия реннин сильно отличается от пепсина, тогда как по структуре близок к нему: так же состоит из одной полипептидной цепи с мол. массой 40000. Изоэлектрическая точка реннина равна 4,5.
Три другие важные эндопептидазы: трипсин, химотрипсин и эластаза, а также одна экзопептидаза — карбоксипептидаза, участвующие в дальнейшем после действия пепсина в переваривании белков, синтезируются в поджелудочной железе. Все они вырабатываются в неактивной форме, в виде проферментов, и их превращение в активные ферменты происходит в тонкой кишке, куда они поступают с панкреатическим соком.
Трипсин. Трипсиноген и трипсин получены в кристаллическом виде, полностью расшифрована их первичная структура и известен молекулярный механизм превращения профермента в активный фермент. В опытах in vitro превращение трипсиногена в трипсин катализируют не только энтеропептидаза и сам трипсин, но и другие протеиназы и ионы Са2+.
Активирование трипсиногена химически выражается в отщеплении с N-конца полипептидной цепи 6 аминокислотных остатков (Вал—Асп—Асп—Асп—Асп—Лиз) и соответственно в укорочении полипептидной цепи (рис. 12.1).
Следует подчеркнуть, что в этом небольшом, казалось бы, химическом процессе — отщепление гексапептида от предшественника — заключено важное биологическое значение, поскольку при этом происходят формирование активного центра и образование трехмерной структуры трипсина, а известно (см. главы 1 и 4), что и белки биологически активны только в своей нативной трехмерной конформации. В том, что трипсин, как и другие протеиназы, вырабатывается в поджелудочной железе в неактивной форме, также имеется определенный физиологический смысл, поскольку в противном случае трипсин мог бы оказывать разрушающее протеолитическое действие не только на клетки самой железы, но и на другие ферменты, синтезируемые в ней (амилаза, липаза и др.). В то же время поджелудочная железа защищает себя еще одним механизмом — синтезом специфического белка ингибитора панкреатического трипсина. Этот ингибитор оказался низкомолекулярным пептидом (мол. масса 6000), который прочно связывается с активными центрами трипсина и химотрипсина, вызывая обратимое их ингибирование. В поджелудочной железе синтезируется также а1-антипротеиназа (мол. масса 50000), которая преимущественно ингибирует эластазу.
* Поступление пищевого белка в желудок стимулирует секрецию гормона гастрина, который в свою очередь стимулирует секрецию НСl и пепсиногена в клетках слизистой оболочки.
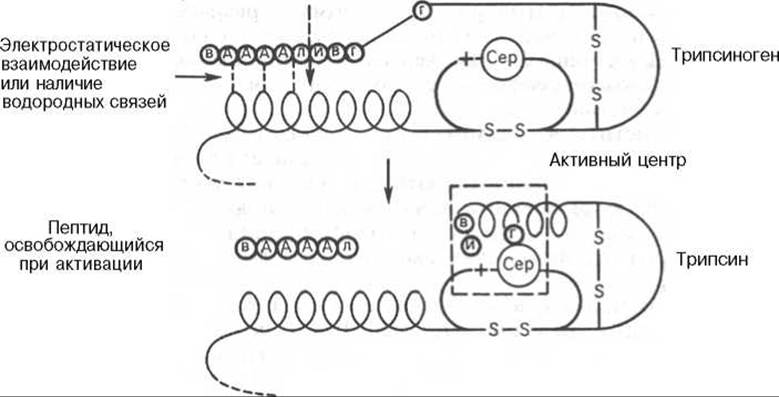
Рис. 12.1. Механизм активации трипсиногена быка (схема).
При остром панкреатите, когда трипсин и другие ферменты из пораженной поджелудочной железы «вымываются» в кровь, уровень их в крови соответствует размерам некротического участка. В этом случае определение активности трипсина в сыворотке крови является надежным ферментным тестом при диагностике острого панкреатита. Следует отметить, что субстратная специфичность трипсина ограничена разрывом только тех пептидных связей, в образовании которых участвуют карбоксильные группы лизина и аргинина.
Химотрипсин. В поджелудочной железе синтезируется ряд химотрипсинов (a-, ß- и л-химотрипсины) из двух предшественников — химотрипсиногена А и химотрипсиногена В. Активируются проферменты в кишечнике под действием активного трипсина и химотрипсина. Полностью раскрыта последовательность аминокислот химотрипсиногена А, во многом сходная с последовательностью аминокислот трипсина. Молекулярная масса его составляет примерно 25000. Он состоит из одной полипептидной цепи, содержащей 246 аминокислотных остатков. Активация профермента не сопряжена с отщеплением большого участка молекулы (см. рис. 4.3). Получены доказательства, что разрыв одной пептидной связи между аргинином и изолейцином в молекуле химотрипсиногена А под действием трипсина приводит к формированию п-химотрипсина, обладающего наибольшей ферментативной активностью. Последующее отщепление дипептида Сер—Арг приводит к образованию δ-химотрипсина. Аутокаталитический процесс активирования, вызванный химотрипсином, сначала способствует формированию неактивного промежуточного неохимотрипсина, который под действием активного трипсина превращается в a-химотрипсин; этот же продукт образуется из δ-химотрипсина, но под действием активного химотрипсина.
Таким образом, благодаря совместному перекрестному воздействию химотрипсина и трипсина из химотрипсиногена образуются разные химотрипсины, различающиеся как ферментативной активностью, так и некоторыми физико-химическими свойствами, в частности электрофоретической подвижностью.
Следует отметить, что химотрипсин обладает более широкой субстратной специфичностью, чем трипсин. Он катализирует гидролиз не только пептидов, но и эфиров, гидроксаматов, амидов и других ацилпроизводных, хотя наибольшую активность химотрипсин проявляет по отношению к пептидным связям, в образовании которых принимают участие карбоксильные группы ароматических аминокислот: фенилаланина, тирозина и триптофана*.
Эластаза. В поджелудочной железе синтезируется еще одна эндопептидаза — эластаза — в виде проэластазы. Превращение профермента в эластазу в тонкой кишке катализируется трипсином. Название фермент получил от субстрата эластина, который он гидролизует. Эластин содержится в соединительной ткани и характеризуется наличием большого числа остатков глицина и серина. Эластаза обладает широкой субстратной специфичностью, но предпочтительнее гидролизует пептидные связи, образованные аминокислотами с небольшими гидрофобными радикалами, в частности глицином, аланином и серином. Интересно, что ни трипсин, ни химотрипсин не гидролизуют пептидные связи молекулы эластина, хотя все три фермента, включая эластазу, содержат сходные участки аминокислотных последовательностей и одинаковые места положения дисульфидных мостиков, а также имеют в активном центре один и тот же ключевой остаток серина (см. табл. 4.2), что подтверждают опыты с ингибированием всех трех ферментов диизопропилфторфосфатом, химически связывающим ОН-группу серина. Высказано предположение, что все три эндопептидазы поджелудочной железы: трипсин, химотрипсин и эластаза, — возможно, имеют один и тот же общий предшественник и что специфичность активного фермента в основном определяется конформационными изменениями профермента в процессе активирования.
Экзопептидазы. В переваривании белков в тонкой кишке активное участие принимает семейство экзопептидаз. Одни из них — карбоксипептидазы — синтезируются в поджелудочной железе в виде прокарбоксипептидазы и активируются трипсином в кишечнике; другие — аминопептидазы — секретируются в клетках слизистой оболочки кишечника и также активируются трипсином.
Карбоксипептидазы. Подробно изучены две карбоксипептидазы — А и В, относящиеся к металлопротеинам и катализирующие отщепление от полипептида С-концевых аминокислот. Карбоксипептидаза А разрывает преимущественно пептидные связи, образованные концевыми ароматическими аминокислотами, а карбоксипептидаза В — связи, в образовании которых участвуют С-концевые лизин и аргинин. Очищенный препарат карбоксипептидазы А обладает бифункциональной активностью — пептидазной и эстеразной и содержит ион Zn2+ (один атом на 1 моль фермента). При замене ионов Zn2+ на ионы Са2+ полностью утрачивается пептидазная активность, но усиливается исходная эстеразная активность, хотя при этом существенных изменений в третичной структуре фермента не отмечается.
* Химотрипсин является одним из наиболее изученных ферментов, для которого в деталях расшифрован механизм ферментативного катализа, включающий образование промежуточного продукта ацилфермента. Доказана существенность для катализа гидроксильной группы серина и непротонированного остатка гистидина в активном центре фермента.
Аминопептидазы. В кишечном соке открыты два фермента — аланин-аминопептидаза, катализирующая преимущественно гидролиз пептидной связи, в образовании которой участвует N-концевой аланин, и лейцинаминопептидаза, не обладающая строгой субстратной специфичностью и гидролизующая пептидные связи, образованные любой N-концевой аминокислотой. Оба фермента осуществляют ступенчатое отщепление аминокислот от N-конца полипептидной цепи.
Дипептидазы. Процесс переваривания пептидов, их расщепление до свободных аминокислот в тонкой кишке завершают дипептидазы. Среди дипептидаз кишечного сока хорошо изучена глицилглицин-дипептидаза, гидролизующая соответствующий дипептид до двух молекул глицина. Известны также две другие дипептидазы: пролил-дипептидаза (пролиназа), катализирующая гидролиз пептидной связи, в образовании которой участвует СООН-группа пролина, и пролин-дипептидаза (пролидаза), гидролизующая дипептиды, в которых азот пролина связан кислотно-амидной связью.
Еще сравнительно недавно протеиназы традиционно связывали только с процессами переваривания. В настоящее время появляется все больше данных о более широкой биологической роли протеолитических ферментов органов и тканей в регуляции ряда вне- и внутриклеточных процессов. Некоторые протеиназы выполняют защитную функцию (свертывание крови, система комплемента, лизис клеток), другие генерируют гормоны, токсины, вазоактивные агенты (ангиотензин, кинины). Ряд протеиназ регулирует образование пищеварительных ферментов, взаимодействие между клетками и клеточными поверхностями, процессы фертилизации (хитин-синтетаза) и дифференциации. Регуляция в большинстве случаев предусматривает превращение неактивного предшественника в активный белок путем отщепления ограниченного числа пептидов. Этот процесс, впервые описанный К. Линдерстрем-Лангом еще в 50-е годы, в последнее время называют ограниченным протеолизом. Значение его очень важно для понимания сущности биологического синтеза в клетках неактивных преи пробелков. Кроме того, этот процесс нашел широкое практическое применение в лабораториях и промышленности. В регуляции действия протеолитических ферментов участвуют также ингибиторы протеиназ белковой природы, открытые не только в поджелудочной железе, но и в плазме крови, курином яйце и т.д.
Отделение панкреатического и кишечного соков регулируется нейрогормональными факторами, которые подробно излагаются в курсе физиологии. Имеются доказательства роли соляной кислоты в качестве пускового механизма выработки в кишечнике особых гормонов. В частности, соляная кислота, попадая в двенадцатиперстную кишку, стимулирует секрецию секретина (см. главу 8); последний, стимулируя секрецию и отделение щелочного панкреатического сока, способствует оттоку желчи. Показано, что секретин быстро исчезает из кровотока, а новые порции его не вырабатываются, поскольку соляная кислота нейтрализуется щелочным панкреатическим соком. Таким образом, благодаря существованию такого механизма, действующего по типу обратной связи, осуществляется регуляция секреции и отделения поджелудочного сока. Поджелудочный сок, полученный при действии секретина, содержит незначительное количество ферментов, но богат бикарбонатами, создающими слабощелочную среду (рН 7,5—8,5), оптимальную для действия пищеварительных ферментов в кишечнике. Вторым гормоном, также синтезирующимся в двенадцатиперстной кишке и регулирующим секрецию поджелудочного сока, является холецистокинин (панкреозимин); он стимулирует отделение сока, богатого ферментами и бедного бикарбонатами.