Биологическая химия - Березов Т. Т., Коровкин Б. Ф. 1998
Биосинтез белка
Регуляция синтеза белка
Основным условием существования любых живых организмов является наличие тонкой, гибкой, согласованно действующей системы регуляции, в которой все элементы тесно связаны друг с другом. В белковом синтезе не только количественный и качественный состав белков, но и время синтеза имеют большое значение. От этого зависит приспособление микроорганизмов к условиям окружающей питательной среды как биологической необходимости или приспособление сложного многоклеточного организма к физиологическим потребностям при изменении внутренних и внешних условий.
Клетки живых организмов обладают способностью синтезировать огромное количество разнообразных белков. Однако они никогда не синтезируют все белки. Количество и разнообразие белков, в частности ферментов, определяются степенью их участия в метаболизме. Более того, интенсивность обмена регулируется скоростью синтеза белка и параллельно контролируется аллостерическим путем (см. главу 4). Таким образом, синтез белка регулируется внешними и внутренними факторами и условиями, которые диктуют клетке синтез такого количества белка и такого набора белков, которые необходимы для выполнения физиологических функций. Все это свидетельствует о весьма сложном, тонком и целесообразном механизме регуляции синтеза белка в клетке.
Общую теорию регуляции синтеза белка разработали французские ученые, лауреаты Нобелевской премии Ф. Жакоб и Ж. Моно. Сущность этой теории сводится к «выключению» или «включению» генов как функционирующих единиц, к возможности или невозможности проявления их способности передавать закодированную в структурных генах ДНК генетическую информацию на синтез специфических белков. Эта теория, доказанная в опытах на бактериях, получила широкое признание, хотя в эукариотических клетках механизмы регуляции синтеза белка, вероятнее всего, являются более сложными (см. далее). У бактерий доказана индукция ферментов (синтез ферментов de novo) при добавлении в питательную среду субстратов этих ферментов. Добавление конечных продуктов реакции, образование которых катализируется этими же ферментами, напротив, вызывает уменьшение количества синтезируемых ферментов. Это последнее явление получило название репрессии синтеза ферментов. Оба явления — индукция и репрессия — взаимосвязаны.
Согласно теории Ф. Жакоба и Ж. Моно, в биосинтезе белка у бактерий участвуют по крайней мере 3 типа генов: структурные гены, ген-регулятор и ген-оператор. Структурные гены определяют первичную структуру синтезируемого белка. Именно эти гены в цепи ДНК являются основой для биосинтеза мРНК, которая затем поступает в рибосому и, как было указано, служит матрицей для биосинтеза белка. Регуляция синтеза белка путем индукции представлена на рис. 14.12.
Синтез мРНК на структурных генах молекулы ДНК непосредственно контролируется определенным участком, называемым геном-оператором. Он служит как бы пусковым механизмом для функционирования структурных генов. Ген-оператор локализован на крайнем отрезке структурного гена или структурных генов, регулируемых им. «Считывание» генетического кода, т.е. формирование мРНК, начинается с промотора — участка ДНК, расположенного рядом с геном-оператором и являющегося точкой инициации для синтеза мРНК, и распространяется последовательно вдоль оператора и структурных генов. Синтезированную молекулу мРНК, кодирующую синтез нескольких разных белков, принято называть полигенным (полицистронным) транскриптом. Координированный одним оператором одиночный ген или группа структурных генов образует оперон.
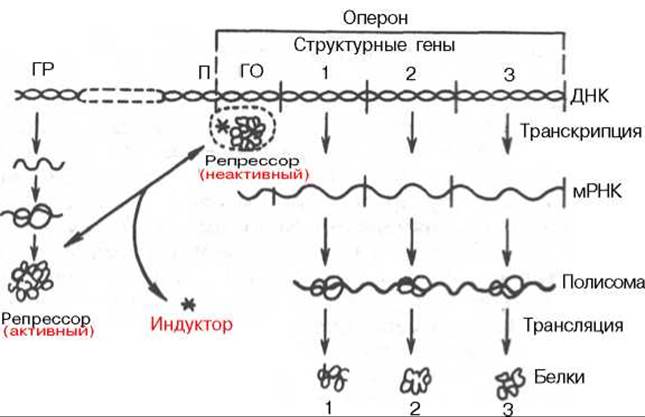
Рис. 14.12. Регуляция синтеза белка путем индукции (схема). ГР - ген-регулятор; П - промотор; ГО - ген-оператор.
В свою очередь деятельность оперона находится под контролирующим влиянием другого участка цепи ДНК, получившего название гена-регулятора. Структурные гены и ген-регулятор расположены в разных участках цепи ДНК, поэтому связь между ними, как предполагают Ф. Жакоб и Ж. Моно, осуществляется при помощи вещества-посредника, оказавшегося белком и названного репрессором. Образование репрессора происходит в рибосомах ядра на матрице специфической мРНК, синтезированной на генерегуляторе (рис. 14.13). Репрессор имеет сродство к гену-оператору и обратимо соединяется с ним в комплекс. Образование такого комплекса приводит к блокированию синтеза мРНК и, следовательно, синтеза белка, т.е. функция гена-регулятора состоит в том, чтобы через белок-репрессор прекращать (запрещать) деятельность структурных генов, синтезирующих мРНК. Репрессор, кроме того, обладает способностью строго специфически связываться с определенными низкомолекулярными веществами, называемыми индукторами, или эффекторами. Если такой индуктор соединяется с репрессором, то последний теряет способность связываться с геном-оператором, который, таким образом, выходит из-под контроля гена-регулятора, и начинается синтез мРНК. Это типичный пример отрицательной формы контроля, когда индуктор, соединяясь с белком-репрессором, вызывает изменения его третичной структуры настолько, что репрессор теряет способность связываться с геном-оператором. Процесс этот аналогичен взаимоотношениям аллостерического центра фермента с эффектором, под влиянием которого изменяется третичная структура фермента и он теряет способность связываться со своим субстратом.
Механизм описанной регуляции синтеза белка и взаимоотношения репрессора со структурными генами были доказаны в опытах с Е. coli на примере синтеза ß-галактозидазы (лактазы) — фермента, расщепляющего молочный сахар на глюкозу и галактозу. Дикий штамм Е. coli обычно растет на глюкозе. Если вместо глюкозы в питательную среду добавить лактозу (новый источник энергии и углерода), то штамм не будет расти, пока не будут синтезированы соответствующие ферменты (адаптивный синтез). При поступлении в клетку лактозы (индуктор) молекулы ее связываются с белком-репрессором и блокируют связь между репрессором и геном-оператором. Ген-оператор и структурные гены при этом начинают снова функционировать и синтезировать необходимую мРНК, которая«дает команду» рибосомам синтезировать ß-галактозидазу. Одновременно ген-регулятор продолжает вырабатывать репрессор, но последний блокируется новыми молекулами лактозы, поэтому синтез фермента продолжается. Как только молекулы лактозы будут полностью расщеплены, репрессор освобождается и, поступив в ДНК, связывает ген-оператор и блокирует синтез мРНК, а следовательно, синтез ß-галактозид азы в рибосомах.
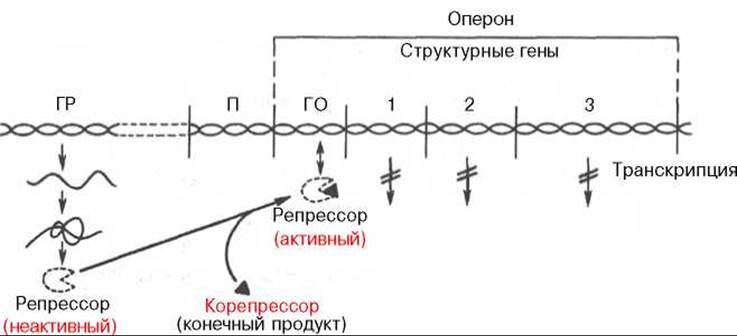
Рис. 14.13. Регуляция синтеза белка путем репрессии (схема). Обозначения те же, что на рис. 14.12.
Таким образом, биосинтез мРНК, контролирующий синтез белка в рибосомах, зависит от функционального состояния репрессора. Этот репрессор представляет собой тетрамерный белок с общей мол. массой около 150000. Если он находится в активном состоянии, т.е. не связан с индуктором, то блокирует ген-оператор и синтеза мРНК не происходит. При поступлении метаболита — индуктора — в клетку его молекулы связывают репрессор, превращая его в неактивную форму (или, возможно, снижают его сродство к гену-оператору). Структурные гены выходят из-под запрещающего контроля и начинают синтезировать нужную мРНК.
Как было указано, концентрация ряда ферментов в клетках резко снижается при повышении содержания отдаленных конечных продуктов, образующихся в цепи последовательных ферментативных реакций. Такой эффект, получивший название репрессии ферментов, часто наблюдается при реакциях биосинтеза. В этих случаях молекулы репрессора, также образующиеся в рибосомах ядра по «команде» гена-регулятора, являются неактивными и сами по себе не обладают способностью подавлять деятельность гена-оператора и, следовательно, всего оперона, но приобретают такую способность после образования комплекса с конечным или одним из конечных продуктов биосинтетического процесса (см. рис. 14.13).
Конечный продукт выступает, таким образом, в качестве корепрессора. Имеются данные, что в качестве корепрессоров в синтезе ферментов обмена аминокислот, по-видимому, выступает не только свободная аминокислота как конечный продукт биосинтетической реакции, но и комплекс ее с тРНК—аминоацил-тРНК.
В регуляции экспрессии структурных генов специфическое участие принимает особый белок — катаболитный генактивирующий белок (от англ. catabolite gene activation protein, сокращенно CAP). Этот белок, взаимодействующий с цАМФ, образует комплекс, способствующий прикреплению РНК-полимеразы к промоторному участку генома. В присутствии комплекса САР-цАМФ фермент может начать транскрипцию оперона, включая структурные гены, т.е. в клетках имеется еще один, дополнительный САР-цАМФ-регулятор, действующий, скорее всего, в качестве положительного регулятора, поскольку его присутствие необходимо для начала экспрессии гена.
Таким образом, концепция Ф. Жакоба и Ж. Моно о механизме проявления (экспрессии) активности генов признана одним из блестящих достижений молекулярной биологии. Она явилась логическим развитием многочисленных исследований, проведенных генетиками и биохимиками в предшествующие десятилетия.
Регуляция экспрессии активности генов у эукариот осуществляется значительно более сложным путем, поскольку процессы транскрипции и трансляции разделены не только пространственно ядерной биомембраной, но и во времени. Эта регуляция базируется как минимум на 6 уровнях сложных биологических процессов, определяющих скорость синтеза и распада генетического продукта (рис. 14.14).
Для большинства эукариотических клеток, как и клеток прокариот, стадия инициации транскрипции является основной, главной регуляторной точкой экспрессии активности генов. Тем не менее имеются существенные различия: во-первых, место процессов транскрипции (в ядре) и трансляции (в цитоплазме); во-вторых, активирование транскрипции у эукариот связано с множеством сложных изменений структуры хроматина в транскрибируемой области; в-третьих, в эукариотических клетках превалируют положительные регуляторные механизмы над отрицательными.
Положительная или отрицательная регуляция определяется типом белков, вовлеченных в механизм регуляции. Получены доказательства существования минимум 3 типов белков, участвующих в регуляции процесса инициации транскрипции, опосредованного через РНК-полимеразу: специфические факторы, репрессоры и активаторы. Первые вызывают изменение специфичности РНК-полимеразы к данному промотору или группе промоторов; репрессоры связываются с промотором, блокируя тем самым доступ РНК-полимеразы к промотору; активаторы, напротив, связываются вблизи промоторного участка, повышая связывание промотора и РНК-полимеразы.
В многоклеточных организмах среднее число регуляторных сайтов для одного гена минимум равно пяти; положительные регуляторные белки связываются со своими специфическими последовательностями в структуре ДНК (вероятнее всего, посредством водородных связей между амидной группой Глн или Асн и пуриновыми и пиримидиновыми основаниями нуклеотидов). Следует указать еще на один момент, почему эукариотическая клетка использует положительные механизмы регуляции экспрессии генов. Подсчитано, что в геноме человека содержится около 100000 генов, соответственно каждая клетка при отрицательном механизме регуляции могла бы синтезировать 100000 разных репрессоров, причем в достаточных количествах. При положительном механизме регуляции большинство генов в принципе неактивно, соответственно молекула РНК-полимеразы не связывается с промотором и клетка синтезирует ограниченный и избирательный круг активаторных белков, необходимых для инициации транскрипции.
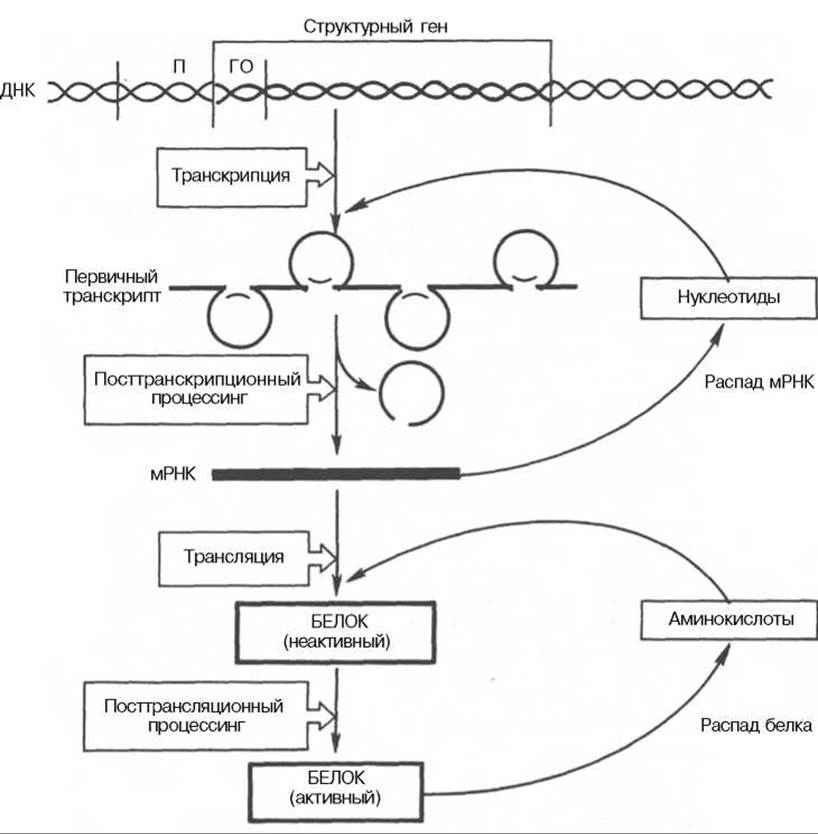
Рис. 14.14. Схематическое изображение регуляции экспрессии активности гена у эукариот.
У эукариот выделены и охарактеризованы также пять регуляторных белков, получивших название транскрипционных факторов (TF: IIА, IIВ, IID, IIЕ и IIF). Они необходимы для узнавания участка (сайта) ДНК, названного TATA (concensus последовательности, ТАТАААА). Детальный молекулярный механизм действия факторов транскрипции пока не раскрыт.
Более подробно в структурном и функциональном отношении у эукариот изучена группа белков, получивших название белков — активаторов транскрипции. Эти белки имеют специфические структурные домены для связывания с другими, но определенными регуляторными нуклеотидными последовательностями в молекуле ДНК. В частности, они содержат домен, специфически связывающийся с ДНК, и один или несколько доменов, необходимых для активирования или взаимодействия с другими регуляторными белками. Среди этих белков — активаторов транскрипции имеются белки, содержащие богатые глутамином домены (до 25%) и богатые пролином домены. Следует отметить, однако, что некоторые из них или почти все регуляторные белки активируют транскрипцию не прямо, а опосредованно — через промежуточные белки, названные коактиваторами. Происхождение и механизм действия последних также не выяснены.
Современные знания о механизмах регуляции экспрессии генов на посттранскрипционном и посттрансляционном уровнях (см. рис. 14.4) были подробно рассмотрены ранее (см. главы 13 и 14).
Рассмотрим кратко вопрос о регуляции процессов дифференцировки клеток высших организмов. ДНК, присутствующая во всех соматических клетках, вероятнее всего, имеет одинаковую первичную структуру у данного организма и соответственно располагает информацией для синтеза любых или всех белков тела. Тем не менее клетки печени, например, синтезируют сывороточные белки, а клетки молочной железы — белки молока. Нет сомнения в том, что в дифференцированных клетках имеется весьма тонкий механизм контроля деятельности ДНК в разных тканях, обеспечивающий синтез многообразия белков.
Механизмы, лежащие в основе этой регуляции, пока неизвестны. Для их объяснения существует ряд гипотез. Предполагают, что контроль осуществляется на уровне транскрипции по аналогии с индукцией ферментов у бактерий и что в этом случае в клетках животных должны функционировать аналогичные репрессоры. С молекулой ДНК у эукариот связаны гистоны, поэтому считается, что именно эти белки выполняют роль репрессоров. Прямых доказательств их роли в качестве репрессоров не получено, хотя, как было показано, в клетках эукариот открыт класс регуляторных белков процесса транскрипции. Высказано предположение, что в ядре синтезируется высокомолекулярная молекула мРНК, содержащая информацию для синтеза широкого разнообразия белков, но в цитоплазму попадает только небольшая часть зрелой мРНК, а основная часть ее распадается. Неясны, однако, биологический смысл и назначение этого механизма избирательного распада и соответственно траты огромной массы молекулы мРНК.
Существует еще одно предположение, что на ДНК клетки синтезируются все мыслимые, возможные мРНК, которые поступают в цитоплазму, и процесс трансляции регулируется путем специфического и избирательного взаимодействия рибосом с определенными молекулами мРНК.