Биологическая химия - Березов Т. Т., Коровкин Б. Ф. 1998
Нервная ткань
Химические основы возникновения и проведения нервных импульсов
Роль медиаторов в передаче нервных импульсов
Связь миллиардов нейронов мозга осуществляется посредством медиаторов. Химическое вещество можно отнести к числу медиаторов лишь в том случае, если оно удовлетворяет ряду критериев. В нервных волокнах должны содержаться ферменты, необходимые для синтеза этого вещества. При раздражении нервов это вещество должно выделяться, реагировать со специфическим рецептором на постсинаптической клетке и вызывать биологическую реакцию. Должны существовать механизмы, быстро прекращающие действие этого вещества.
Всем этим критериям удовлетворяют два вещества — ацетилхолин и норадреналин. Содержащие их нервы называют соответственно холинергическими и адренергическими. В соответствии с этим все эфферентные системы делят на холинорецепторы и адренорецепторы.
Ряд других химических веществ удовлетворяют многим, но не всем перечисленным критериям. К таким медиаторам относят дофамин, адреналин, серотонин, октопамин, гистамин, ГАМК и др.
Обширная группа холинорецепторов весьма неоднородна как в структурном, так и в функциональном отношении. Объединяют их медиатор ацетилхолин и общая схема строения синапса.
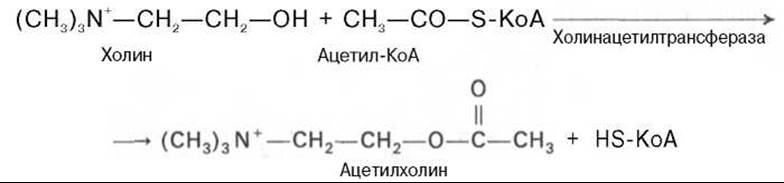
Ацетилхолин* представляет собой сложный эфир уксусной кислоты и холина. Он синтезируется в нервной клетке из холина и активной формы ацетата—ацетилкоэнзима А при помощи специального фермента холин- ацетилтрансферазы (холинацетилазы):
Синапс можно представить себе как узкое пространство (щель), ограниченное с одной стороны пресинаптической, а с другой — постсинаптической мембраной (рис. 19.4). Пресинаптическая мембрана состоит из внутреннего слоя, принадлежащего цитоплазме нервного окончания, и наружного слоя, образованного нейроглией. Мембрана в некоторых местах утолщена и уплотнена, в других истончена и имеет отверстия для сообщения цитоплазмы аксона с синаптическим пространством. Постсинаптическая мембрана менее плотная, не имеет отверстий. Подобным образом построены и нервно-мышечные синапсы, но они имеют более сложное строение мембранного комплекса.
В общих чертах картину участия ацетилхолина в осуществлении передачи нервного импульса возбуждения можно представить следующим образом. В синаптических нервных окончаниях имеются пузырьки (везикулы) диаметром 30—80 нм, которые содержат нейромедиаторы. Эти пузырьки покрыты оболочкой, которая образована белком клатрином (мол. масса 180000). В холинергических синапсах каждый пузырек диаметром 80 нм содержит ~ 40000 молекул ацетилхолина. При возбуждении высвобождение медиатора происходит «квантами», т.е. путем полного опорожнения каждого отдельного пузырька. В нормальных условиях под влиянием сильного импульса выделяется примерно 100—200 квантов медиатора — количество, достаточное для инициирования потенциала действия в постсинаптическом нейроне. Происходит это, по-видимому, следующим образом. Деполяризация мембраны синаптических окончаний вызывает быстрый ток ионов Са2+ в клетку. Временное увеличение внутриклеточной концентрации ионов Са2+ стимулирует слияние мембраны синаптических пузырьков с плазматической мембраной и таким образом запускает процесс высвобождения их содержимого. Для выброса содержимого одного пузырька требуется примерно 4 иона Са2+. Выделенный в синаптическую щель ацетилхолин вступает во взаимодействие с белком-хеморецептором, входящим в состав постсинаптической мембраны. В результате изменяется проницаемость мембраны — резко увеличивается ее пропускная способность для ионов Na+. Взаимодействие между рецептором и медиатором запускает ряд реакций, заставляющих постсинаптическую нервную клетку или эффекторную клетку выполнять свою специфическую функцию. После выделения медиатора должна наступить фаза его быстрой инактивации, или удаления, чтобы подготовить синапс к восприятию нового импульса.
* Ацетилхолин служит также медиатором в двигательных концевых пластинках (нервномышечные соединения, являющиеся участками контактов между нервом и поперечнополосатой мышцей).
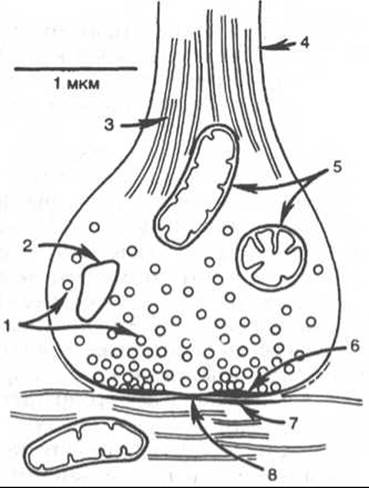
Рис. 19.4. Схематическое изображение синапса (по Мецлеру).
1 - синаптические пузырьки; 2 - лизосома; 3 - микрофибриллы (нейрофибриллы); 4 - аксон; 5 - митохондрии; 6 - пресинаптическое утолщение мембраны; 7 - постсинаптическое утолщение мембраны; 8 - синаптическая щель (около 20 нм).
В холинергических синапсах это происходит двумя путями. Первый путь заключается в том, что ацетилхолин подвергается ферментативному гидролизу. Второй путь — это энергозависимый активный транспорт ацетилхолина в нейрон, где он накапливается для последующего повторного использования.
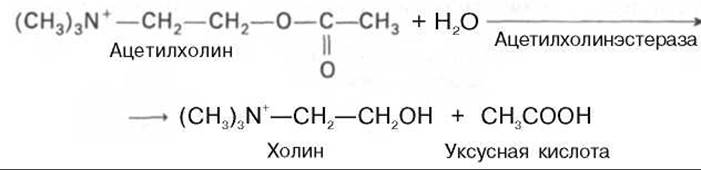
Гидролитический распад ацетилхолина на уксусную кислоту и холин катализируется ферментом, который получил название «ацетилхолинэстераза»:
В большинстве отделов головного мозга гидролиз ацетилхолина осуществляется ацетилхолинэстеразой (истинная холинэстераза, которая гидролизует ацетилхолин быстрее, чем иные эфиры холина). В нервной ткани существуют и другие эстеразы, которые способны гидролизовать ацетилхолин, но значительно медленнее, чем, например, бутирилхолин. Эти эстеразы называются холинэстеразой (или псевдохолинэстеразой). К числу холинергических систем относятся моторные нейроны, образующие нервно-мышечные соединения, все преганглионарные нейроны автономной нервной системы и постганглионарные нейроны парасимпатической нервной системы. Большое количество холинергических симпатических областей обнаружено также в головном мозге. В зависимости от чувствительности к той или иной группе химических соединений холинергические нейроны делятся на мускариновые (активируемые мускарином) и никотиновые (активируемые никотином). Мускариновые рецепторы ацетилхолина, имеющиеся во многих нейронах автономной нервной системы, специфически блокируются атропином. Никотиновые синапсы присутствуют в ганглиях и скелетных мышцах. Их ингибиторами являются кураре и активный компонент этого яда D-тубокурарин.
Необходимо подчеркнуть, что в адренорецепторах существует два вида рецепторов для норадреналина: а- и ß-адренергические рецепторы. Эти рецепторы можно отличить друг от друга по специфическим реакциям, которые они вызывают, а также по тем специфическим агентам, которые способны блокировать данные реакции.
ß-Адренергические рецепторы включают эфферентную клетку с помощью аденозин-3',5'-монофосфата, или цАМФ — универсального «второго посредника» между гормонами и различными функциями клеток, на которые воздействуют гормоны (см. главу 8).
Установлено, что как только ß-адренергический рецептор, расположенный на наружной поверхности мембраны эффекторной клетки, начинает взаимодействовать с норадреналином, на внутренней поверхности клеточной мембраны активируется фермент аденилатциклаза. Затем в клетке аденилатциклаза превращает АТФ в цАМФ; последний в свою очередь способен оказывать влияние на метаболизм клетки. Этот сложный ряд последовательных реакций может быть заблокирован пропранололом — веществом, препятствующим связыванию норадреналина с ß-адренергическим рецептором.
Известно, что в метаболизме катехоламиновых медиаторов особая роль принадлежит ферменту моноаминоксидазе (МАО). Этот фермент удаляет аминогруппу (—NH2) у норадреналина, серотонина, дофамина и адреналина, тем самым инактивируя указанные медиаторы. В последние годы было показано, что, помимо ферментативного превращения, существует и другой механизм быстрой инактивации, точнее удаления, медиаторов. Оказалось, что норадреналин быстро исчезает из синаптической щели в результате вторичного поглощения симпатическими нервами; вновь оказавшись в нервном волокне, медиатор, естественно, не может воздействовать на постсинаптические клетки. Конкретный механизм этого явления пока не вполне ясен.
Адренергическая и холинергическая системы головного мозга тесно взаимодействуют с другими системами мозга, в частности использующими серотонин в качестве медиатора. В основном серотонинсодержащие нейроны сосредоточены в ядрах мозгового ствола. Нейромедиаторная роль серотонина осуществляется в результате взаимодействия серотонина со специфическими серотонинергическими рецепторами. Исследования, проведенные с ингибитором синтеза серотонина n-хлорфенилаланином, а также с другими ингибиторами, дают основания считать, что серотонин влияет на процессы сна. Выявлено также, что торможение кортикостероидами секреторной активности гипофиза оказывается менее эффективным у тех животных, мозг которых беднее серотонином.
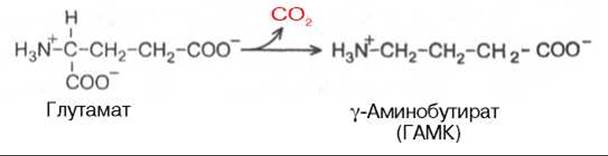
Важным нейромедиатором, выполняющим тормозные функции, является у-аминомасляная кислота (ГАМК), количество которой в головном мозге во много раз больше, чем других нейромедиаторов. Так, в гипоталамусе суммарное содержание ацетилхолина, норадреналина, дофамина и серотонина не превышает 10 мкг/г, в то время как ГАМК в этом отделе головного мозга более 600 мкг/г. ГАМК увеличивает проницаемость постсинаптических мембран для ионов К+ и тем самым отдаляет мембранный потенциал от порогового уровня, при котором возникает потенциал действия; таким образом, ГАМК — это тормозной нейромедиатор. ГАМК образуется при декарбоксилировании глутамата в реакции, катализируемой глутаматдекарбоксилазой:
В терапевтической практике применяется большое количество лекарственных средств, которые действуют через систему медиаторов. Многие лекарственные препараты, успешно применяемые при лечении гипертонии, влияют на накопление и выделение адренергических медиаторов. Например, резерпин — понижающее артериальное давление средство специфически тормозит процесс переноса катехоламинов в специальные гранулы нейронов и тем самым делает эти амины доступными действию эндогенной МАО.
Гипотензивные лекарственные препараты, такие, как а-метилдофа, под действием содержащихся в нервной клетке (аксоне) ферментов превращаются в вещества, напоминающие по своему строению норадреналин. Эти «ложные» медиаторы накапливаются и выделяются вместе с естественными медиаторами, «разбавляя» их и тем самым снижая их эффект.
Многие антидепрессанты (вещества, снимающие депрессию) увеличивают содержание катехоламинов в синаптической щели, т.е. количество медиаторов для стимулирования рецептора возрастает. К таким веществам, в частности, относятся имипрамин (блокирует поглощение норадреналина нервными волокнами), амфетамин (одновременно способствует выделению норадреналина и блокирует его поглощение), ингибиторы МАО (подавляют метаболизм катехоламинов) и др. В связи с этим возникла катехол-аминовая гипотеза депрессивных состояний, согласно которой психическая депрессия связана с недостатком катехоламинов в мозге.
В начале 50-х годов фармакологи выяснили, что известный галлюциноген диэтиламин лизергиновой кислоты (ЛСД) не только сходен по химическому строению с серотонином, но и нейтрализует некоторые его фармакологические эффекты (блокируя рецепторы серотонина). Поэтому было высказано предположение, что нарушение обмена серотонина может быть причиной возникновения особых психических заболеваний.
Считают, что такие антипсихотические средства, как аминазин (хлорпромазин) и галоперидол, усиливая синтез катехоламинов, способны блокировать дофаминовые рецепторы в мозге.