Биологическая химия - Березов Т. Т., Коровкин Б. Ф. 1998
Химия нуклеиновых кислот
Структура нуклеиновых кислот
Третичная структура нуклеиновых кислот
Выделить нативную молекулу ДНК (рис. 3.2) из большинства источников, в частности хромосом, чрезвычайно трудно из-за высокой чувствительности молекулы ДНК к нуклеазам тканей и гидродинамической деструкции*.
Удалось выделить в интактном (неповрежденном) виде только некоторые ДНК вирусов, митохондрий и хлоропластов. Исследования этих молекул при помощи физических (в частности, кристаллографических) и физико-химических методов показали, что двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации с образованием суперспирали или открытой кольцевой формы. Оказалось также, что линейная ДНК может образоваться из кольцевой формы или существовать как таковая в природе. В некоторых вирусах обнаружены, кроме того, одноцепочечные ДНК линейной и кольцевой форм (рис. 3.3).
Образование кольцевой формы молекулы ДНК у бактерий или в митохондриях клеток животных часто вызвано ковалентным соединением их открытых концов. Известно, что суперспиральная (суперскрученная) структура обеспечивает экономную упаковку огромной молекулы ДНК в хромосоме: вместо 8 см длины, которую она могла бы иметь в вытянутой форме, в хромосоме человека молекула ДНК настолько плотно упакована, что ее длина составляет 5 нм. Обычно в ДНК встречаются положительные и отрицательные супервитки, образованные за счет скручивания по часовой (правосторонней) или против часовой стрелки двойной спирали. Образование подобных супервитков катализируется специфическими ферментами, получившими название топоизомераз. Подобные суперспирали соединяются с белками (гистонами), упакованными в бороздках, обеспечивая тем самым стабильность третичной структуры ДНК. Степень суперспиральности (наличие супервитков) молекулы ДНК обычно устанавливают по изменению константы седиментации в определенных условиях. Суперспирализация ДНК может быть нарушена разрывом в одной из цепей или в обеих цепях двойной спирали под действием ДНКазы или при обработке интеркалирующими соединениями. Под интеркаляцией подразумевают встраивание плоских ароматических колец между стопками пар азотистых оснований ДНК. Интеркаляция может быть вызвана антибиотиками и красителями; в интактных клетках она может быть обусловлена ароматическими кольцами амнокислот, что имеет, очевидно, определенный биологический смысл в проблеме белково-нуклеинового узнавания.
* Разработан щадящий метод выделения нативной молекулы ДНК с использованием протеиназы К и ДСН. Выделенный из клеток почек обезьяны препарат ДНК имел необычно высокую мол. массу — 2 ∙ 108, однако даже в этом случае молекулярная масса на несколько порядков меньше, чем мол. масса ДНК in vivo, исчисляемая 1010—1011.

Рис. 3.2. Модель молекулы ДНК.
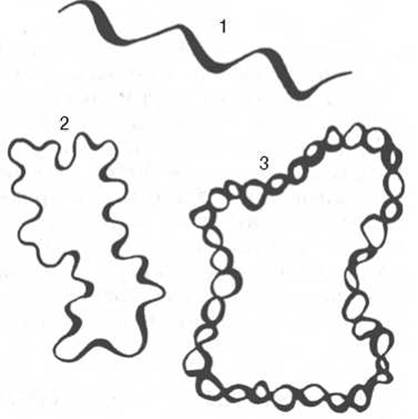
Рис. 3.3. Третичная структура ДНК (схема).
1 - линейная одноцепочечная ДНК - бактериофаг фХ174 и другие вирусы; 2 - кольцевая одноцепочечная ДНК вирусов и митохондрий; 3 - кольцевая двойная спираль ДНК.
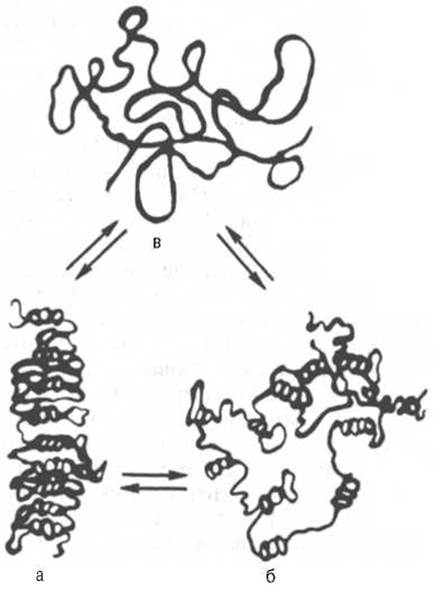
Рис. 3.4. Третичная структура РНК в растворе в зависимости от ионной силы, температуры и рН среды (схема) (по А. С. Спирину и Л.П. Гавриловой).
а - компактная палочка, б - компактный клубок; в - развернутая цепь.
Данные о структуре тРНК свидетельствуют о том, что нативные молекулы тРНК имеют примерно одинаковую третичную структуру, которая отличается от плоской структуры «клеверного листа» большой компактностью за счет складывания различных частей молекулы. Следует указать на существование у ряда вирусов (реовирус, вирус раневых опухолей растений и др.) природных двухцепочечных РНК, обладающих однотипной с ДНК структурой. При физиологических значениях рН среды, ионной силы и температуры создаются условия для образования в одноцепочечных матричных и рибосомных РНК множества участков с двойной спиралью («шпильки») и дальнейшего формирования комплементарных участков, определяющих в известной степени жесткость их третичной структуры (рис. 3.4). В настоящее время получены доказательства значимости ван-дер-ваальсовых (диполь-дипольных и лондоновских) связей между азотистыми основаниями в стабилизации общей пространственной конфигурации нуклеиновых кислот.