БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ - Е. С. Северина - 2004
РАЗДЕЛ 4. БИОСИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТ И БЕЛКОВ (МАТРИЧНЫЕ БИОСИНТЕЗЫ). ОСНОВЫ МОЛЕКУЛЯРНОЙ ГЕНЕТИКИ
VII. Регуляция экспрессии генов у про- и эукариотов
Организмы адаптируются к меняющимся условиям окружающей среды путём изменения экспрессии (скорости транскрипции) генов. Этот процесс, в деталях изученный на бактериях и вирусах, включает взаимодействие специфических белков с участками ДНК в непосредственной близости от стартового участка транскрипции. При этом может происходить включение или выключение транскрипции. Эукариотические клетки используют тот же самый принцип, хотя в регуляции реализуются и некоторые другие более сложные механизмы.
А. Регуляция активности генов у прокариотов. Теория оперона
Исследования на клетках Е. coli позволили установить, что у бактерий существуют ферменты 3 типов: конститутивные, присутствующие в клетках в постоянных количествах независимо от метаболического состояния организма (например, ферменты гликолиза); индуцируемые, их концентрация в обычных условиях мала, но может возрастать в 1000 раз и более, если, например, в среду культивирования клеток добавить субстрат такого фермента; репрессируемые, т. е. ферменты метаболических путей, синтез которых прекращается при добавлении в среду выращивания конечного продукта этих путей.
1. Теория оперона
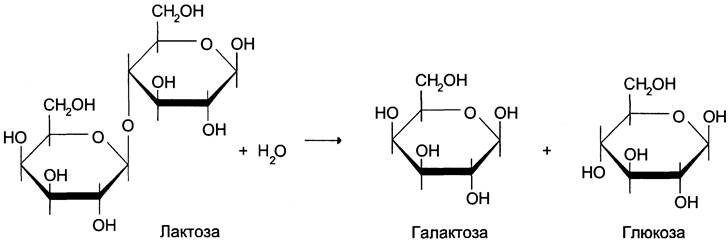
На основании генетических исследований индукции ß-галактозидазы, участвующей в клетках Е. coli, в гидролитическом расщеплении лактозы (рис. 4-46), Франсуа Жакоб и Жак Моно в 1961 г. сформулировали гипотезу оперона, которая объясняла механизм контроля синтеза белков у прокариотов.
Рис. 4-46. Гидролиз лактозы β-галактозидазой.
В экспериментах гипотеза оперона получила полное подтверждение, а предложенный в ней тип регуляции стали называть контролем синтеза белка на уровне транскрипции, так как в этом случае изменение скорости синтеза белков осуществляется за счёт изменения скорости транскрипции генов, т. е. на стадии образования мРНК.
У Е. coli, как и у других прокариотов, ДНК не отделена от цитоплазмы ядерной оболочкой. В процессе транскрипции образуются первичные транскрипты, не содержащие интронов, а мРНК лишены «кэпа» и поли-А-конца. Синтез белка начинается до того, как заканчивается синтез его матрицы, т. е. транскрипция и трансляция протекают почти одновременно. Исходя из размера генома (4 x 106 пар нуклеотидов), каждая клетка Е. coli содержит информацию о нескольких тысячах белков. Но при нормальных условиях роста она синтезирует около 600-800 различных белков, а это означает, что многие гены не транскрибируются, т. е. неактивны. Гены белков, функции которых в метаболических процессах тесно связаны, часто в геноме группируются вместе в структурные единицы (опероны). Согласно теории Жакоба и Моно, оперонами называют участки молекулы ДНК, которые содержат информацию о группе функционально взаимосвязанных структурных белков, и регуляторную зону, контролирующую транскрипцию этих генов. Структурные гены оперона экспрессируются согласованно, либо все они транскрибируются, и тогда оперон активен, либо ни один из генов не «прочитывается», и тогда оперон неактивен. Когда оперон активен и все его гены транскрибируются, то синтезируется полицистронная мРНК, служащая матрицей для синтеза всех белков этого оперона. Транскрипция структурных генов зависит от способности РНК-полимеразы присоединяться к промотору, расположенному на 5'-конце оперона перед структурными генами.
Связывание РНК-полимеразы с промотором зависит от присутствия белка-репрессора на смежном с промотором участке, который называют «оператор». Белок-репрессор синтезируется в клетке с постоянной скоростью и имеет сродство к операторному участку. Структурно участки промотора и оператора частично перекрываются, поэтому присоединение белка-репрессора к оператору создаёт стерическое препятствие для присоединения РНК-полимеразы.
Большинство механизмов регуляции синтеза белков направлено на изменение скорости связывания РНК-полимеразы с промотором, влияя таким образом на этап инициации транскрипции. Гены, осуществляющие синтез регуляторных белков, могут быть удалены от оперона, транскрипцию которого они контролируют.
2. Индукция синтеза белков. Lac-оперон
Теория оперона была предложена на основании данных, полученных при изучении свойств лактозного оперона (lас-оперона) Е. coli, т. е. оперона, в котором закодированы белки, участвующие в усвоении лактозы.
Клетки Е. coli обычно растут на среде, используя в качестве источника углерода глюкозу. Если в среде культивирования глюкозу заменить на дисахарид лактозу, то по прошествии нескольких минут клетки адаптируются к изменившимся условиям. Они начинают продуцировать 3 белка, обеспечивающих утилизацию лактозы. Один из этих белков — фермент β-галактозидаза, катализирующий гидролитическое расщепление лактозы до глюкозы и галактозы.
В присутствии глюкозы клетки Е. соli содержат менее 10 молекул этих ферментов на клетку. Перенос клеток на среду, содержащую лактозу, вызывает индукцию — увеличение количества молекул каждого из ферментов до 5000 (рис. 4-47).
Рис. 4-47. Механизм индукции лактозного оперона. А - в отсутствие индуктора (лактозы) белок-репрессор связан с оператором. РНК-полимераза не может присоединиться к промотору, транскрипция структурных генов оперона не идёт; Б - в присутствии лактозы белок-репрессор присоединяет её, изменяет свою конформацию и теряет сродство к оператору. РНК-полимераза связывается с промотором и транскрибирует структурные гены: β-галактозидазы (А), катализирующей гидролиз лактозы до глюкозы и галактозы; галактозидпермеазы (В), осуществляющей транспорт лактозы и других галактозидов в клетки; тиогалактозидтрансацетилазы (С) — фермента, способного переносить ацетильную группу ацетил-КоА на тиогалактозу. Функция его в процессе утилизации лактозы пока неясна.
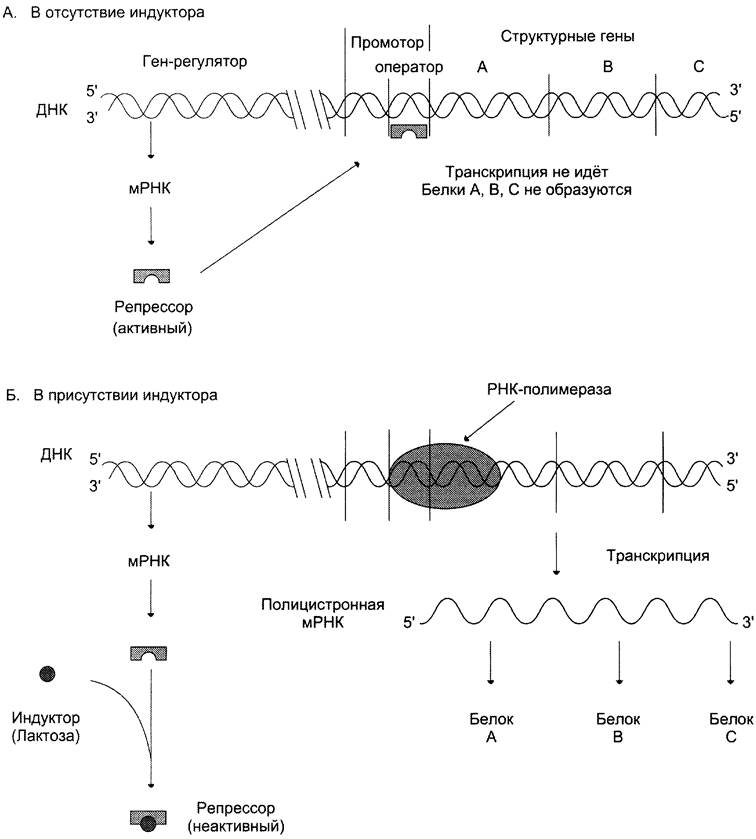
Теория оперона объясняет это явление следующим образом. В отсутствие индуктора (лактозы) белок-репрессор связан с оператором. А поскольку участки оператора и промотора перекрываются, то присоединение репрессора к оператору препятствует связыванию РНК-полимеразы с промотором, и транскрипция структурных генов оперона не идёт. Когда в среде появляется индуктор, т. е. лактоза, то он присоединяется к белку-репрессору, изменяет его конформацию и снижает сродство к оператору. РНК-полимераза связывается с промотором и транскрибирует структурные гены.
3. Репрессия синтеза белков. Триптофановый и гистидиновый опероны
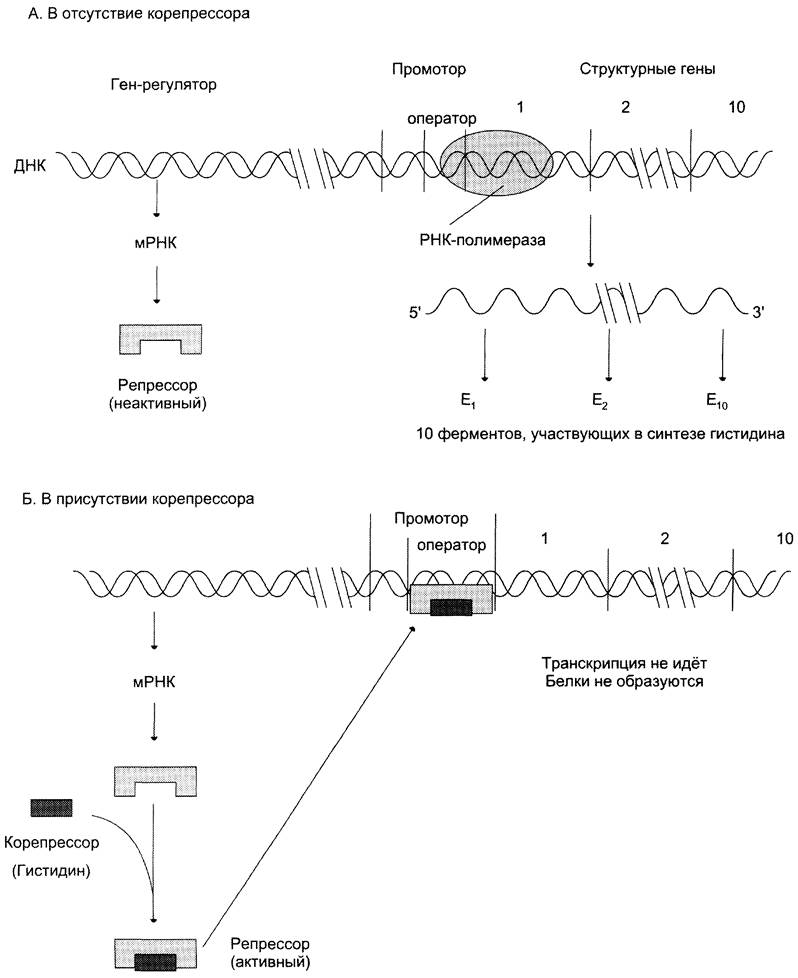
Снижение концентрации фермента в бактериальной клетке может осуществляться путём репрессии синтеза ферментов. Сущность этого механизма регуляции заключается в следующем: когда клетки Е. соИ растут на среде, содержащей в качестве единственного источника азота соль аммония, то им приходится синтезировать все азотсодержащие вещества. Такие клетки, в частности, должны содержать все ферменты, необходимые для синтеза 20 различных аминокислот. Однако если добавить в среду культивирования одну из аминокислот, например, триптофан или гистидин, то клетка перестанет вырабатывать весь набор ферментов, необходимых для синтеза этих аминокислот из аммиака и источника углерода. Репрессия синтеза ферментов, катализирующих последовательность реакций метаболического пути конечным продуктом, как это имеет место в случае ферментов синтеза гистидина или триптофана, называется репрессией конечным продуктом.
Это явление теория оперона объясняет следующим образом: при отсутствии в среде Гис или Три регуляторный белок-репрессор не имеет сродства к оператору и происходит синтез ферментов, осуществляющих образование этих аминокислот. Когда в среду добавляют, например, Гис, то эта небольшая молекула, получившая название «корепрессор»,присоединяется к белку-репрессору. В результате конформационных изменений в молекуле репрессора комплекс белка-репрессора и корепрессора (Гис) приобретает сродство к оператору, присоединяется к нему, и транскрипция оперона прекращается, т. е. прекращается считывание информации о строении 10 ферментов, участвующих в синтезе этой аминокислоты (рис. 4-48).
Рис. 4-48. Механизм репрессии синтеза ферментов, участвующих в образовании гистидина. А - в отсутствие корепрессо- ра (гистидина) белок-репрессор не имеет сродства к оператору, РНК-полимераза присоединяется к промотору, и происходит транскрипция 10 структурных генов, кодирующих строение ферментов, участвующих в синтезе гистидина; Б - в присутствии гистидина в среде комплекс белка-репрессора с корепрессором, т. е. Гис, связывается с оператором, препятствует присоединению РНК-полимеразы к промотору и останавливает транскрипцию.
Следует иметь в виду, что репрессия и индукция синтеза белков у прокариотов реализуют принципы адаптации к меняющимся условиям существования и клеточной экономии: ферменты появляются в клетках, когда в них существует потребность, и перестают вырабатываться, если потребность исчезает.
Б. Механизмы регуляции экспрессии генов у эукариотов
Эукариотические организмы (и особенно млекопитающие) устроены значительно сложнее прокариотов и нуждаются в более сложном аппарате регуляции. Так, в организме человека имеется более 200 различных типов клеток, существенно различающихся по структуре и функциям. В то же время различными методами исследования ДНК (прежде всего, методом молекулярной гибридизации) доказано, что количество и структура ДНК практически всех клеток организма одинаковы (за исключением лимфоцитов), т. е. все клетки организма содержат один и тот же геном. У высших организмов по сравнению с прокариотическими существенно возрастает содержание ДНК на гаплоидную клетку: с 4,2х 106 пар нуклеотидов у Е. coli до 3,3 х 109 пар нуклеотидов в клетках человека.
1. Организация хроматина в дифференцированных клетках многоклеточного организма
В клетках млекопитающих наряду с адаптивной регуляцией, обеспечивающей приспособление организма к меняющимся условиям внутренней и внешней среды, существуют механизмы, которые сохраняют стабильную (существующую на протяжении всей жизни клетки и даже многих её генераций) репрессию одних генов и депрессию других.
В ядрах дифференцированных клеток хроматин имеет такую укладку, что только небольшое число генов (часто менее 1%) доступно для транскрипции. Различают участки гетерохроматина, в которых ДНК упакована очень компактно и недоступна для транскрипции, и участки эухроматина, имеющие более рыхлую укладку и способные связывать РНК-полимеразу. В разных типах клеток в область эухроматина попадают разные гены, а это означает, что в разных тканях транскрибируются разные участки хроматина.
Стойкая репрессия генов гетерохроматина обеспечивается:
✵ пространственной укладкой ДНК, при которой гетерохроматин находится в высоко- конденсированном состоянии;
✵ метилированием дезоксицитидина ДНК-метилазами в 5'-CG-3' последовательностях ДНК. Эта модификация сильно меняет конформацию хроматина и препятствует активной транскрипции;
✵ связыванием с гистонами и образованием нуклеосом, которые также снижают транскрипционную активность ДНК.
Исследования показали, что области эухроматина, в которых расположены активно транскрибируемые гены, обладают некоторыми структурными особенностями:
✵ они более чувствительны к действию ДНКаз, чем остальные участки ДНК;
✵ молекулы гистонов, связанные с ДНК в этих участках, модифицированы: ε-аминогруппа лизина метилирована или ацетилирована; метилированы некоторые остатки аргинина и гистидина в гистонах Н2А и Н2В, являющихся коровыми белками нуклеосом. Некоторые молекулы Н2А образуют прочный комплекс с белком убиквитином. В гистоне Н1 фосфорилируются остатки серина. Результат этой серии ковалентных модификаций — снижение суммарного, положительного заряда гистонов и ослабление сродства нуклеосом к ДНК.
✵ к областям «активного» хроматина присоединяется группа негистоновых НMG-белков, или белков с высокой подвижностью при гель-электрофорезе. Эти белки содержат много положительно заряженных аминокислотных остатков, связывание с которыми ослабляет взаимодействие ДНК и гистонов и вызывает дополнительное повышение транскрипционной активности генов.
Разнообразие клеток и возросшая сложность клеточных процессов нуждаются в большом разнообразии механизмов регуляции. Показано, что разный набор и количество белков в эукариотических клетках может регулироваться:
✵ изменением количества структурных генов;
✵ перестройкой генов в хромосомах;
✵ эффективностью транскрипции разных участков генома;
✵ характером посттранскрипционных модификаций первичных транскриптов;
✵ на уровне трансляции;
✵ с помощью посттрансляционных превращений вновь синтезированных полипептидных цепей.
2. Изменение количества генов
Геном эукариотов обнаруживает высокую пластичность, играющую важную роль в регуляции активности некоторых генов и увеличивающую разнообразие клеточных ответов. У млекопитающих реализуются следующие варианты изменений в структуре генов:
Амплификация (или увеличение числа) генов используется организмом в том случае, когда возникает необходимость увеличить синтез определённого генного продукта. Многие гены, кодирующие белки или РНК, необходимые организму в больших количествах (например, гистоны, рРНК, тРНК), постоянно присутствуют в амплифицированном состоянии. Так, у человека 20% общего генома состоит из участков, кодирующих рибосомные, транспортные и ядрышковые РНК, последние из которых обеспечивают посттранскрипционные модификации РНК. Амплифицированные участки могут располагаться друг за другом (тандемно) в хромосоме или образовывать внехромосомные фрагменты ДНК, называемые двойными мини-хромосомами, их размер колеблется от 100 до 1000 килобаз (1 килобаза = 1000 пар нуклеотидов). Описано более 20 генов, способных амплифицироваться при определённых условиях.
К числу генов, для которых обнаружена амплификация, относят ген металлотионеина. Продукт экспрессии этого гена — низкомолекулярный белок металлотионеин, обладающий способностью связывать тяжёлые металлы (медь, цинк, кадмий, ртуть) и защищать клетки от отравления этими соединениями. Установлено, что в ответ на повышение концентрации тяжёлых металлов в крови в клетках происходит амплификация гена металлотионеина.
Другими примерами генов, количество которых увеличивается под влиянием лекарственных препаратов, являются ген дищдрофолатредукгазы (см. разделы 9, 10) и ген Р-гликопротеина, ответственный за синтез белка, обеспечивающего множественную лекарственную устойчивость опухолевых клеток (см. раздел 16).
Утрата генетического материала — довольно редкий способ регуляции. Наиболее яркий пример потери всех генов за счёт разрушения ядра — процесс созревания эритроцитов. Нестабильны амплифицированные гены, двойные хромосомы. Они, как правило, исчезают в последующих генерациях. Утрата генетического материала происходит в процессе созревания лимфоцитов и образования плазматических клеток разных клонов, синтезирующих секретируемые формы иммуноглобулинов.
3. Перестройка генов
У высших организмов, так же, как и у прокариотов, отмечают процесс обмена, перемещения генов между хромосомами или внутри хромосомы, объединение генов с образованием изменённой хромосомы, которая после таких структурных изменений способна к репликации и транскрипции. Этот процесс получил название «генетическая рекомбинация».
У эукариотов рекомбинации наблюдают:
✵ при половом слиянии яйцеклетки и сперматозоида;
✵ при перемещении подвижных генетических элементов — транспозонов, в состав которых входят отдельные гены или группа генов, с исходной позиции в какое-либо другое место той же или другой хромосомы;
✵ при формировании в лимфоцитах «библиотеки» генов, кодирующих антитела или иммуноглобулины.
Рассмотрим более подробно механизмы, обеспечивающие образование в организме каждого человека около 10 млн (107) различных антител, т.е. количества значительно большего, чем число всех других белков, существующих у каждого индивидуума. Антитела с одинаковыми антигенсвязывающими свойствами синтезируются В-лимфоцитами, принадлежащими к одному определённому клону (т.е. группе клеток, возникшей из одной родоначальной клетки). При попадании в организм любого антигена среди имеющегося набора В-лимфоцитов всегда найдётся такой клон клеток, антитела которого имеют комплементарный ему активный центр. Антитела встроены в плазматическую мембрану В-лимфоцитов, и их антигенсвязывающие участки локализованы на поверхности клеток. Антиген, присоединяясь к активному центру антитела, вызывает пролиферацию клеток и превращение В-лимфоцитов в плазматические клетки, в которых идут активный синтез и секреция не связанных с мембраной антител.
Изучение вопроса о происхождении антител позволило сделать вывод о том, что огромное многообразие белков иммунной системы кодируется ограниченным количеством генетического материала, изменения в котором обеспечиваются рекомбинациями и соматическими мутациями (или изменениями в структуре ДНК,
которые сохраняются при последующих делениях клеток).
Вспомним, что мономерные антитела — доменные белки, состоящие из двух идентичных тяжёлых (Н) цепей и двух идентичных лёгких (L) цепей. Лёгкие цепи имеют двухдоменную структуру и включают вариабельный (VL) и константный (СL) домены. Тяжёлая цепь состоит из 4 — 5 доменов: одного вариабельного (VH) и, как правило, трёх константных (СH). Иммуноглобулины — гликопротеины; их углеводная часть присоединяется к константной области Н-цепей. В связывании антигенов участвуют 2 активных центра антитела, образованные вариабельными областями Н-цепей (VH) и L-цепей (VL). L-цепи бывают двух типов: λ (лямбда) и к κ (каппа), значительно различающиеся по первичной структуре С-областей.
Наличие в антителах С- и V-областей позволило предположить, что гены, обеспечивающие синтез L- и Н-цепей, образуются в результате соединения двух участков гена, один из которых кодирует вариабельную область, а второй — константную. И действительно вскоре было установлено, что в зародышевых клетках и соматических клетках, не синтезирующих иммуноглобулины, участки гена, кодирующие V- и С-области L-цепей λ-типа, разделены протяжёнными нуклеотидными последовательностями, но сближены в зрелых В-лимфоцитах, синтезирующих Lλ. Из этого следовал вывод о том, что в процессе дифференцировки В-лимфоцита из зародышевой клетки происходит «вырезание» протяжённого участка генетического материала, обеспечивающее сближение VL- и CL-областей с образованием полного гена L-цепи иммуноглобулина. Этот процесс перестройки в геноме получил название соматической рекомбинации, так как он связан с созреванием лимфоцитов и не передаётся по наследству.
Описано 3 разных семейства генных фрагментов, или сегментов, кодирующих строение L- и Н-цепей Ig. Два семейства ответственны за синтез лёгких цепей: сегменты, кодирующие строение L-цепей типа λ, расположены в хромосоме 22, генетический материал L-цепей типа к (каппа) — в хромосоме 2, а информация о всём разнообразии Н-цепей локализована в хромосоме 14.
Полные гены L-цепей λ и к типов в ходе дифференцировки собираются из 3 сегментов: вариабельного (VL), соединительного (JL) и константного (СL). Так, для L-цепей к типа обнаружено около 300 сегментов, кодирующих N-концевой вариабельный (Vк) участок полипептидной цепи длиной в 95 аминокислотных остатков, 5 сегментов, в которых содержится информация об остальных 13 аминокислотах Vк области, и 1 сегмент константной области. В зародышевых клетках 300 сегментов Ук расположены в хромосоме последовательно на расстоянии 7 килобаз друг от друга. Каждый V-сегмент состоит из 2 экзонов, разделённых коротким интроном: лидирующий экзон (L) кодирует сигнальный пептид (20 — 25 аминокислотных остатков), а экзон Vк — основную часть вариабельного домена. Семейство Vк-сегментов отделено от группы соединительных сегментов (Jк) участком ДНК размером в 20 килобаз. Между последним из Jк-сегментов и Ск-экзоном, кодирующим домен константной области, расположен интрон размером в 2,4 килобазы (рис. 4-49).
Рис. 4-49. Образование гена лёгкой цепи к (каппа) типа и его транскрипция. 1 — \/-сегменты L-цепей состоят из двух экзонов и разделяющего их интрона. Экзоны кодируют сигнальный пептид (L) и почти весь вариабельный домен V. В ходе соматической рекомбинации сегмент гена вариабельной части L-цепи (V2) объединяется с одним из соединительных сегментов (J4); 2 — транскрибируется полный ген L-цепи, состоящий из трёх экзонов и двух интронов, расположенных в следующем порядке: 5’L-I1-, V2J4-J5-I2-C-3'; 3 — в ходе сплайсинга первичного транскрипта гена интрон I2 сегмента V2, J5 сегмент и интрон I2, отделяющий V2J4 от С- области, удаляются.
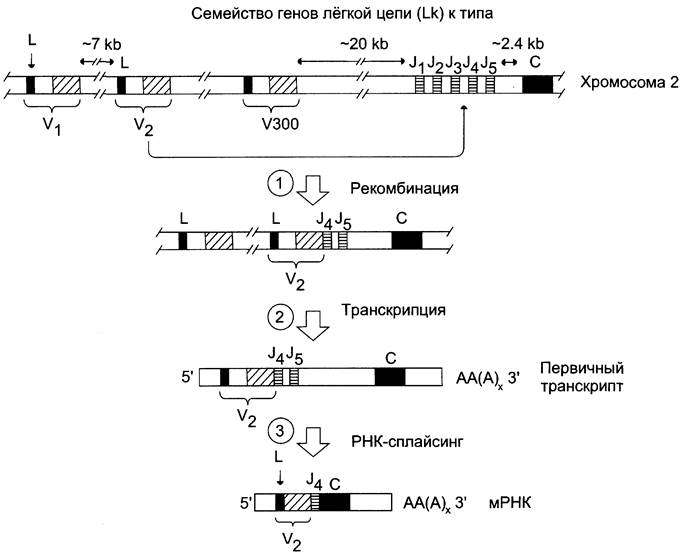
В ходе дифференцировки В-клеток один из вариабельных VL-сегментов путём соматической рекомбинации переносится из отдалённого участка в участок той же хромосомы, рядом с одним из сегментов JL. Например, сегмент V2 объединяется с соединительным мини-сегментом J4, и формируется полный ген L-цепи. Он состоит из 3 экзонов и 2 интронов, расположенных в гене в следующем порядке: 5'L-I1-V2-J4-J5-I2- С-3', где L — лидерная последовательность, кодирующая сигнальный пептид, I1 — интрон V2- сегмента, а I2 — интрон между семейством JL- сегментов и Ск-экзоном. После транскрипции гена в ходе сплайсинга из первичного транскрипта удаляются интроны I1, I2 и лишний сегмент J5, а все кодирующие последовательности соединяются в единую информационную молекулу зрелой мРНК. В процессе синтеза L-цепи на рибосоме лидерный участок, состоящий в основном из гидрофобных аминокислот, обеспечивает прохождение белка через мембрану ЭР и затем отщепляется. Образуется L-цепь, имеющая аминокислотный состав, характерный для L-цепи к-типа в молекуле Ig.
Расчёты показывают, что из имеющихся сегментов к-гена в организме можно синтезировать 4500 полных генов, кодирующих L-цепи к-типа.
Формирование полных генов L-цепей λ-типа происходит так же, как L-цепей к-типа.
Ещё большее разнообразие вариантов возникает при сборке полных генов тяжёлых (Н) цепей Ig. Н-цепи кодируются четырьмя сегментами: VH, DH (от англ. diversity) сегментами разнообразия, JH и СH. У человека обнаружено около 500 VH, 15 DH и 4 JH сегментов.
Каждый VH сегмент содержит информацию об аминокислотной последовательности сигнального пептида и около 100 аминокислот VH домена. В сегменте DH закодирован участок полипептидной цепи, содержащий от 2 до 13 аминокислот, а в сегменте JH — 4 — 6 аминокислот. Полный ген вариабельного домена образуется путём соединения VH, DH и JH сегментов.
При формировании полного гена вариабельной части Н-цепи Ig, состоящей из VH, D и JH сегментов, происходит 2 рекомбинационные состыковки: на первом этапе удаляется участок между выбранными Dx и Jy кодирующими последовательностями, а на втором — между Vi и DxJy сегментами. Экзонов, кодирующих константную область Н-цепей, описано 10: Сμ, Сσ, Су3, Су1, Сα1, Су2α, Су2β, Су4, Сε и Сα2, они определяют классы и подклассы иммуноглобулинов — IgM, IgG, IgA и т. д.
Первыми в иммунном ответе появляются IgM, поскольку к полному гену вариабельного домена ближе всех остальных С-экзонов находится Cμ сегмент Н-цепи. Активированные В- клетки могут синтезировать мембранно-связанную и секретируемую формы IgM. Кроме того, они могут переключаться с синтеза IgM на образование антител других классов. Перед каждым СH экзоном имеется участок ДНК, называемый «участок переключения», или «свич-сайт» (от англ. swich site), построенный из повторяющихся нуклеотидных последовательностей. Эти участки облегчают протекание дополнительной рекомбинации, в ходе которой удаляются С-сегменты между полным геном вариабельной области и С-сегментом того класса, который должен быть включён.
Исследование нуклеотидных последовательностей генов некоторых подклассов L-цепей к- типа и Н-цепей показало, что разнообразие структуры сегментов, закодированных в зародышевой клетке, увеличивают соматические мутации. Мутации происходят в дифференцированных клетках на участках VL-JL и VHDHJH
сегментов в процессе или после рекомбинаций, делая, таким образом, количество антител практически неограниченным. Очень важно, что мутации происходят в областях, ответственных за узнавание антигенов, обеспечивая более полное соответствие активного центра антитела антигену.
Таким образом, перестройки генетического материала в процессе формирования полных генов Ig происходят в несколько этапов, каждый из которых приурочен к строго определённой стадии дифференцировки В-лимфоцитов. Из сегментов, которые кодируют различные участки полипептид ной цепи, входящей в вариабельные домены, и одного из экзонов константного домена собираются полные гены тяжёлых и лёгких нитей Ig. Сборка L-цепей включает одну соматическую рекомбинацию, а сборка Н-цепей происходит с помощью двух соматических рекомбинаций. Когда В-лимфоциты синтезируют Ig не класса М, то это сопровождается ещё одним дополнительным рекомбинационным событием. Соматические мутации, происходящие в зрелых В-лимфоцитах, делают многообразие антител неисчерпаемым.
Аналогичные процессы наблюдают и в ходе дифференцировки Т-лимфоцитов.
4. Регуляция транскрипции
Регуляция транскрипции генов высших организмов сходна с регуляцией экспрессии генов прокариотов. Основное различие состоит в значительно большем количестве участков ДНК и регуляторных факторов, контролирующих этот процесс.
У животных и человека различные гены экспрессируются в разные моменты времени и с разной интенсивностью. Здесь, так же, как у прокариотов, есть гены «домашнего хозяйства», транскрибирующиеся конститутивно, т. е. постоянно и во всех тканях. Это гены гликолиза, синтеза РНК и некоторых белков (например, альбумина). Существуют гены, транскрибирующиеся только в специализированных клетках, т. е. имеет место тканеспецифическая экспрессия. Например, экспрессия генов α- и β-цепей глобина происходит только в клетках-предшественниках эритроцитов. Многие гены подвергаются адаптивной регуляции и являются объектами индуцибельных воздействий или негативного контроля.
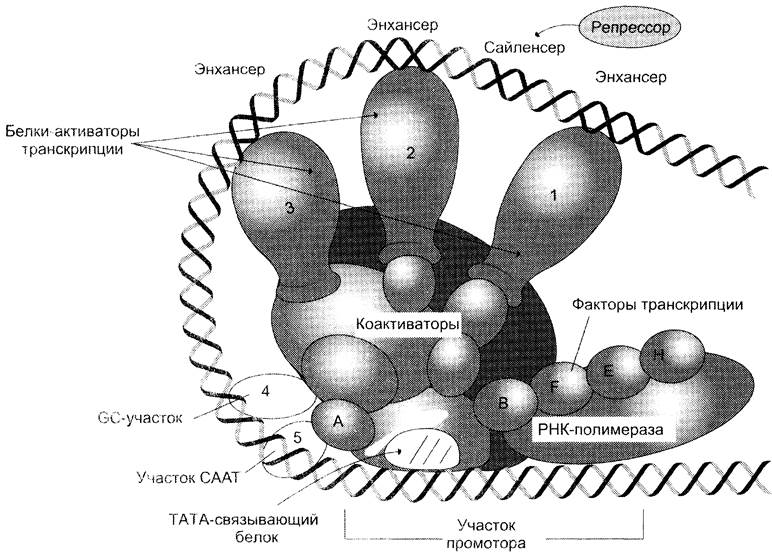
Ранее уже говорилось о том, что минимальный синтез любого белка поддерживается в том случае, если к ТАТА-участку промотора присоединяется ТАТА-связывающий белок, факторы транскрипции и РНК-полимераза, образующие инициирующий комплекс, осуществляющий синтез небольшого количества мРНК. Формирование комплекса — многоступенчатый процесс, от образования которого зависит скорость инициации транскрипции. Идентифицировано более 100 различных белков, способных взаимодействовать со специфическими регуляторными последовательностями ДНК, влияя главным образом на процесс сборки транскрипционного комплекса и скорость транскрипции (рис. 4-50).
Рис. 4-50. Адаптивней регуляция транскрипции у эукариотов. Промоторы эукариотических генов находятся под контролем большого числа регуляторных участков на молекуле ДНК: ТАТА-, СААТ-, GC-последовательностей, энхансеров, сайленсеров- последовательностей, к которым присоединяются комплексы белков с различными лигандами (цАМФ, стероидными гормонами, метаболитами, ионами металлов и т. д.).
Эти белки имеют один или несколько доменов, обеспечивающих выполнение регуляторных функций.
ДНК-связывающие домены, ответственные за узнавание и связывание регуляторных факторов со специфическими участками на молекуле ДНК;
Домены, активирующие транскрипцию за счёт связывания с белками основного инициаторного комплекса: транскрипционными факторами, коактиваторами и РНК-полимеразой;
Антирепрессорные домены, благодаря которым белки способны взаимодействовать с гистонами нуклеосом и освобождать транскрибируемые участки ДНК от связи с этими ингибиторными структурами;
Домены, связывающие лиганды, присоединение которых к белку изменяет его конформацию и обеспечивает связывание с молекулой ДНК. Лиганды-индукторы транскрипции — стероидные гормоны, ретиноевая кислота, кальцитриол (производное витамина D3) и гормоны щитовидной железы. Лигандами-репрессорами могут быть конечные продукты метаболических путей, некоторые гормоны. Будучи липофильными молекулами, они проходят плазматическую, а иногда и ядерную мембраны, взаимодействуют с внутриклеточными рецепторами, присоединяясь к лиганд-связывающему участку (рис. 4-51).
Рис. 4-51. Действие лиганда-индуктора транскрипции на клетку млекопитающих. Лиганд-индуктор. например стероидный гормон связывается с внутриклеточным рецептором, находящимся в ядре или цитоплазме, и поступает в ядро. Комплекс гормон-рецептор присоединяется к определённому участку на молекуле ДНК и активирует транскрипцию гена. Образуется мРНК - матрица для синтеза белка, обеспечивающего определённый клеточный ответ.
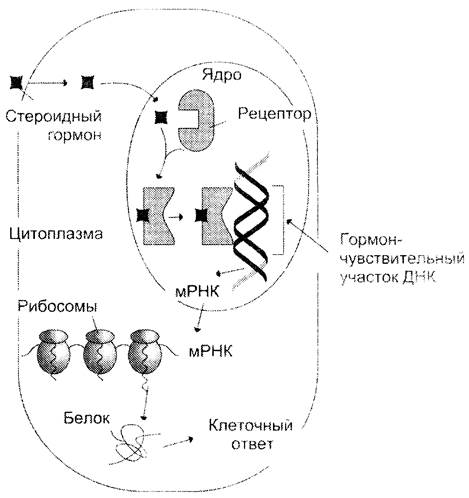
Присоединение лиганда к рецептору образует ДНК-связывающий участок, узнающий специфическую последовательность в регуляторной зоне ДНК и индуцирующий транскрипцию определённых генов.
На молекуле ДНК на расстоянии 100-200 пар оснований от стартовой точки транскрипции имеются короткие специфические последовательности ДНК: СААТ — элемент (или бокс), СG-бокс и октамерный бокс (включающий 8 пар оснований), узнающие транскрипционные факторы. Эти элементы есть во всех клетках, и конститутивно экспрессируемые гены нуждаются только в них. В то же время для генов, подвергающихся адаптивной регуляции, обнаружены участки молекулы ДНК, которые удалены (до 1000 и более нар оснований) от промотора, но тоже участвующие в регуляции транскрипции. Эти нуклеотидные последовательности бывают 2 типов.
Энхансеры — участки ДНК размером 10 — 20 пар оснований, присоединение к которым регуляторных белков увеличивает скорость транскрипции. Если участки ДНК, связываясь с белками, обеспечивают замедление транскрипции, то их называют сайленсерами.
Эти структурные элементы молекулы ДНК контролируют транскрипцию, даже если они:
✵ ориентированы на молекуле ДНК в любом направлении (от 5'- к 3'-концу или наоборот);
✵ связываются с одним иди несколькими регуляторными белками;
✵ располагаются перед или после гена, экспрессию которого они регулируют.
Элементы ответа, или cis-элементы — регуляторные последовательности ДНК, общие для группы генов. Они обеспечивают координированную регуляцию транскрипции генов и, как правило, располагаются на расстоянии примерно в 250 пар оснований выше промогора каждою гена. В остальном эти нуклеотидные последовательности имеют много общего с энхансерами. В данном варианте регуляции один и тог же индуктор, связываясь с соответствующим регуляторным белком, может активировать много разных генов, так как каждый из них в регуляторной области содержит один и тот же cis-элемент. Один из белков-продуктов этой группы генов может оказаться индуктором другой группы генов. Конечный результат регуляции — серия ответных реакций за счёт активации различных генов одним индуктором (рис. 4-52).
Рис. 4-52. Активация группы генов с помощью одного индуктора. Группа генов имеет общий регуляторный cis-элемент и активируется одним и тем же регуляторным белком. Один из белковых продуктов первой серии ответных реакций активирует вторую серию генов (* — cis-элементы к белку X; # — cis-элементы к белку С).
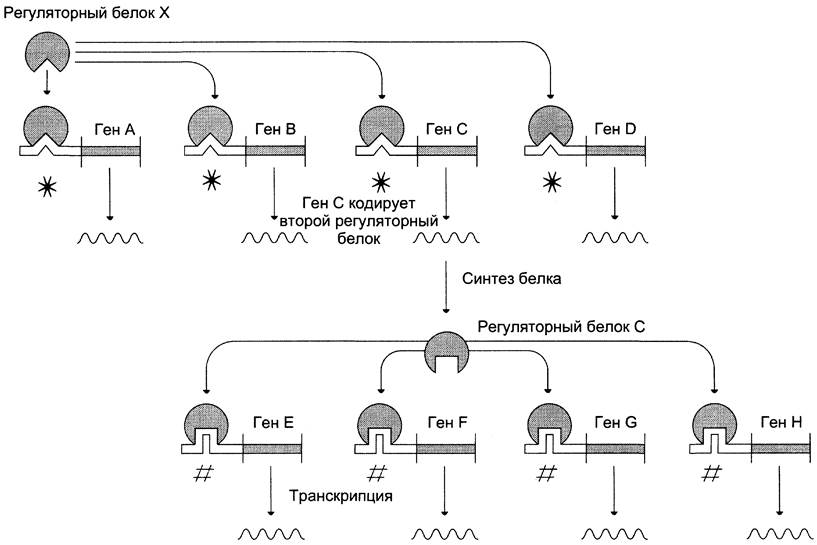
К генам, регулируемым cis-элементами, относят гены, чувствительные к стероидным гормонам, гены белков теплового шока и многие другие. Например, при повышении температуры или после какого-либо другого клеточного стресса активируется синтез транскрипционного фактора, который индуцирует транскрипцию генов, кодирующих строение шаперонов.
Очевидно, что эффективность регуляции во многом зависит от структуры транскрипционных факторов и внутриклеточных рецепторов, непосредственно взаимодействующих с молекулой ДНК. Установлено, что большинство ДНК - связывающих белков принадлежит к трём семействам в зависимости от структуры домена, непосредственно взаимодействующего с двойной спиралью ДНК. Эти белки включают структуры типа «спираль-поворот-спираль», «цинковые пальцы» и «лейциновой молнии» (см. раздел 1). Как правило, эти структуры — небольшие фрагменты молекул белков, а сайт- специфическое связывание происходит за счёт взаимодействия между радикалами аминокислот этих участков и азотистыми основаниями молекулы ДНК.
5. Посттранскрипционная регуляция
В организме животных существенное значение в обеспечении разнообразия белков играет посттранскрипционный процессинг РНК. Основные способы такой регуляции — альтернативный сплайсинг и изменение стабильности РНК.
Альтернативный сплайсинг. Установлено, что многие эукариотические гены, будучи транскрибированы, образуют несколько вариантов зрелой мРНК в ходе процессинга (или созревания) первичного транскрипта, имеющего полиэкзонное строение.
Возможные варианты сплайсинга РНК представлены на рис. 4-53.
Рис. 4-53. Часто встречающиеся варианты сплайсинга первичных транскриптов РНК. I. Вырезание одного из экзонов: а) синтез белка, содержащего полный набор экзонов (1-5); б) синтез белка, лишённого одного экзона (1,2, 4, 5); II. Сохранение участка интрона: а) с 5'-конца; б) с 3'-конца. III. Сохранение целого интрона. IV. Использование альтернативных промоторов (либо перед экзоном 1, либо перед экзоном 2). V. Использование альтернативных участков полиаденилирования (например, при последовательном сшивании экзонов после экзона 3, а если экзон 3 не прочитывается, то после экзона 4).
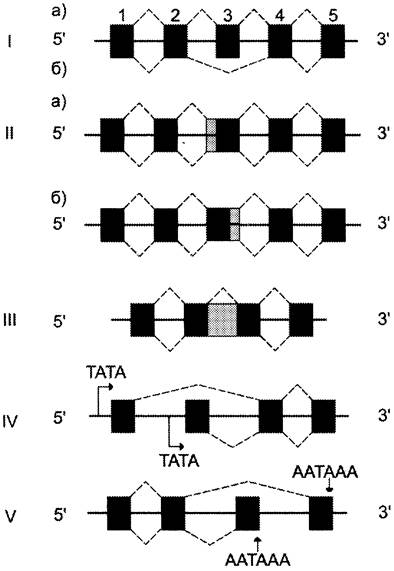
Наиболее часто промотор сохраняется на одном из концов транскрипта, а в ходе сплайсинга происходит «вырезание» одного или нескольких экзонов. В других случаях в зрелой мРНК сохраняется часть интрона и включается в состав экзона с 5' или 3'-конца. Сплайсинг может влиять на выбор промотора или участка полиаденилирования.
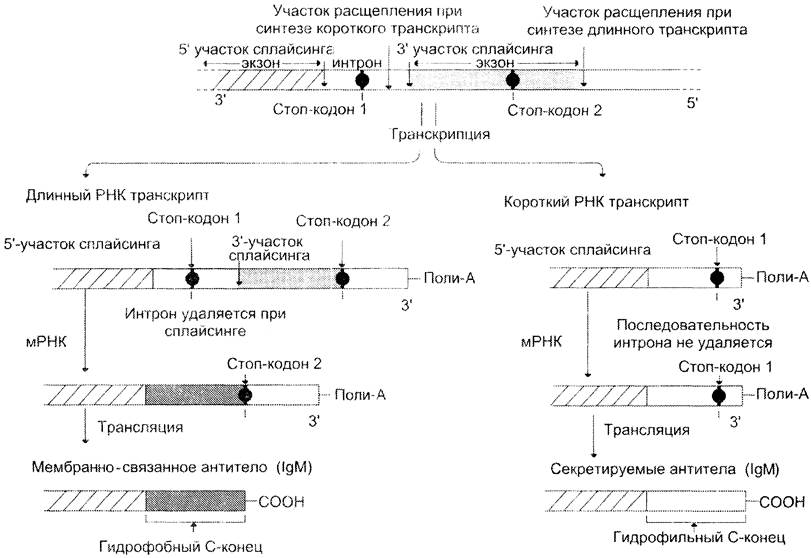
С помощью альтернативного сплайсинга в процессе синтеза антител образуются мембраносвязанные и секреторные формы антител (рис. 4-54). Так, первоначально В-лимфоциты продуцируют транскрипты, полиаденилированные после второго стоп-кодона, а нитрон, в котором имеется первый стоп-кодон, удаляется. В результате синтезируются IgM, связанные с клеточной мембраной, так как мРНК таких клеток содержит на 3'-конце экзон, кодирующий участок полипептидной цепи, состоящий из гидрофобных аминокислот. С помощью этого участка происходит «заякоривание» IgM в мембране. Когда В-лимфоциты превращаются в плазматические клетки, то в
результате альтернативного сплайсинга образуется мРНК, в которой сохраняется интрон, содержащий первый стоп-кодон. Поэтому происходит более раннее полиаденилирование и исчезает экзон, кодирующий гидрофобный участок молекулы. Синтезируются укороченные молекулы антител, секретируемые в кровь.
Рис. 4-54. Использование механизмов альтернативного сплайсинга и полиаденилирования в ходе синтеза мембранно- связанных и секреторных Ig. Если транскрипт подвергается полиаденилированию после второго стоп-кодона, присутствующего в экзоне гена IgM, то синтезируются белки, у которых на С-конце присутствует гидрофобный домен, обеспечивающий связывание с плазматической мембраной. При стимуляции В-лимфоцитов в клетках осуществляется альтернативный сплайсинг первичного транскрипта, при котором интрон, содержащий первый стоп-кодон, сохраняется. Образуются более короткие мРHК, полиаденилирование которых происходит после первого стоп-кодона.
«Редактирование» РНК. Описан ряд случаев, когда первичная структура мРНК изменяется («редактируется») после транскрипции. Последовательность нуклеотидов в таких генах одинакова, а транскрибируемая в разных тканях мРНК различается в результате появления в молекуле замен, вставок или выпадений нуклеотидов. Пример «редактирования» РНК — образование апопротеина В (апо-В) в клетках печени и тонкого кишечника (рис. 4-55). Апо-В — основной компонент липопротеинов, участвующих в транспорте триацилглицеринов из этих тканей в кровь. Хотя апопротеин В кодируется одним и тем же геном, вариант белка, образующийся в печени, называют апо-В-100, и он содержит 4563 аминокислотных остатка, тогда как белок, синтезированный в клетках кишечника, состоит из 2152 аминокислот. В гене, кодирующем этот белок, последовательность нуклеотидов в триплете 2153 — CAA и шифрует включение в полипептидную цепь остатка глутамина. В клетках кишечника в первичном транскрипте гена азотистое основание — цитозин (С) кодона 2153 дезаминируется и превращается в урацил (U). Возникает стоп-кодон — UAA, прекращающий трансляцию мРНК в середине молекулы и приводящий к синтезу укороченного белка. В результате образуется белок (В-48), длина которого составляет 48% от длины белка, синтезируемого печенью.
Рис. 4-55. «Редактирование» мРНК апопротеина В. В ходе транскрипции гена апопротеина В в печени образуется мРНК, служащая матрицей для синтеза белка, состоящего из 4563 аминокислотных остатков. В клетках тонкого кишечника экспрессия того же гена вызывает образование белка, состоящего из 2152 аминокислот. В РНК транскрипте цитозин кодона 2153 — САА превращается в урацил (II), и возникает стоп-кодон в середине молекулы мРНК. Это приводит к синтезу укороченного белка.
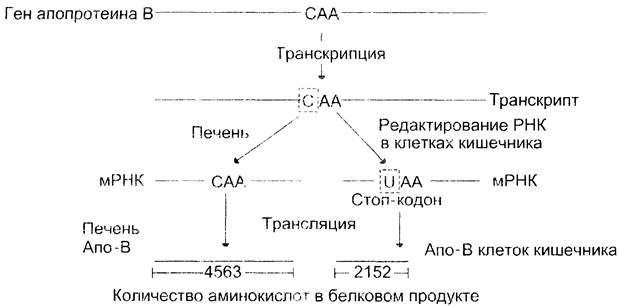
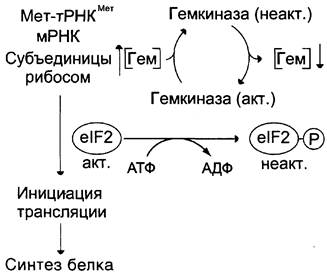
Рисунок 4-56. Зависимость скорости синтеза глобина от концентрации гема. Когда внутриклеточный уровень гема высок, фактор инициации еIF2 не фосфорилирован и активен, происходит синтез глобина. Если содержание гема в клетке снижается, фактор инициации фосфорилируется, инактивируется и синтез белка прекращается.
Изменение стабильности мРНК. Для того, чтобы участвовать в синтезе белка, мРНК должна выйти из ядра в цитоплазму через ядерные поры. Установлено, что в ядре клеток обычно синтезируется больший набор гетерогенных РНК, чем тот, что выходит в цитоплазму. Многие продукты транскрипции подвергаются расщеплению нуклеазами, а те мРНК, что, транспортируются из ядра в цитоплазму, защищаются от гидролитического разрушения, образуя комплексы с белками.
Время жизни эукариотических мРНК значительно больше (t1/2составляет от нескольких часов до нескольких дней), чем t1/2 мРНК прокариотов, равное нескольким минутам. Очевидно, что стабильность молекул мРНК — фактор, изменение которого влияет на уровень трансляции. Стабилизация мРНК при фиксированной скорости транскрипции приводит к накоплению и увеличению количества образующегося белкового продукта.
Продолжительность жизни разных мРНК варьирует в достаточно широких пределах. Некоторые гены кодируют продукт с большой продолжительностью жизни. Так, в ходе транскрипции гена β-глобина образуется мРНК с t1/2, равной примерно 10 ч. Другие гены образуют мРНК с короткой продолжительностью жизни: мРНК, на которых синтезируются факторы роста, имеют t1/2, менее 1 ч. Показано, что поли(А)- фрагмент на 3'-конце мРНК увеличивает продолжительность жизни молекул. Чем длиннее поли(А)-фрагмент, тем больше время жизни мРНК.
Описано много примеров регуляции количества синтезирующихся белков за счёт изменения продолжительности функционирования мРНК. Так, стабильность мРНК-матриц для синтеза молекул гистонов сильно зависит от фазы клеточного цикла. В S-фазе гистоны постоянно синтезируются и используются для укладки, вновь образованной ДНК в нуклеосомы. Гистоновая мРНК в этот период стабильна в течение нескольких часов. После S-периода, когда ДНК уже не синтезируется, в клетках образуется небольшое количество гистонов, так как они не требуются для формирования нуклеосом. В этот период t1/2 для гистоновой мРНК составляет 10-15 мин.
6. Регуляция трансляции и посттрансляционных модификаций
Изменение скорости трансляции
Хотя изменение скорости образования белков на уровне трансляции не относят к числу основных способов регуляции количества и разнообразия белков, некоторые случаи такой регуляции известны. Наиболее изученный пример — синтез белков в ретикулоцитах. Известно, что на этом уровне дифференцировки кроветворные клетки лишены ядра, а, следовательно, и ДНК. Регуляция синтеза белка-глобина осуществляется только на уровне трансляции и зависит от содержания гема в клетке (рис. 4-56). Если внутриклеточная концентрация гема высока, то глобин синтезируется; когда содержание гема снижается, то ингибируется и образование глобина. Остановка синтеза белка осуществляется за счёт фосфорилирования фактора инициации еIF2, который в фосфорилированной форме неактивен. Гем предотвращает фосфорилирование еIF2, связываясь со специфической протеинки- назой, которая получила название гемкиназы.
Некоторые мРНК содержат элементы вторичной структуры на 5'- или 3'-концах нетранслируемого участка мРНК, к которым могут присоединяться белки и ингибировать трансляцию. Например, синтез ферритина — белка, обеспечивающего хранение ионов железа в клетке, усиливается при повышении внутриклеточной концентрации железа (см. раздел 14). Обнаружено, что мРНК ферритина на 5'-конце имеет петли, к которым при низкой концентрации железа присоединяется регуляторный белок. Когда этот белок связан с мРНК, то трансляция не идёт. Если концентрация ионов железа в клетке повышается, то Fе3+ взаимодействует с белком, изменяет его конформацию и сродство к мРНК. мРНК освобождается от регуляторного белка, и на ней начинается синтез ферритина.
Различия в продолжительности жизни молекул белка
После того как белки синтезированы, время их жизни регулируется протеазами. Разные белки имеют разные t1/2: от нескольких часов до нескольких месяцев, а иногда и лет (табл. 4-6). В каждой клетке скорость расщепления белков варьирует в широких пределах. Ферменты, катализирующие регуляторные реакции метаболических путей, как правило, подвергаются быстрому расщеплению, поэтому скорость обновления этих молекул достаточно высока. Физиологическое состояние организма также влияет на продолжительность жизни белков. Кроме того, существует мощная система защиты, обеспечивающая быстрое расщепление дефектных белков.
Таблица 4-6. Период полураспада некоторых белков в клетках млекопитающих
Фермент |
Т1/2. ч |
Орнитиндекарбоксилаза |
0,5 |
Тирозинаминотрансфераза |
2,0 |
Карбоксикиназа фосфоенолпирувата |
5,0 |
Аргиназа |
96 |
Альдолаза |
118 |
Лактатдегидрогеназа |
144 |
Цитохром С |
150 |
Некоторые белки расщепляются лизосомными ферментами. В процессе аутофагии содержимое клетки, включая органеллы, окружается мембраной, сливается с л изосомой другой клетки и подвергается действию лизосомных ферментов. В результате гидролиза образующиеся мономеры поступают в цитоплазму для повторного использования.
Для других белков показано расщепление в цитоплазме протеазами. Так, подлежащие разрушению белки первоначально отмечаются клеткой путём присоединения белка под названием убиквитин. Этот небольшой белок, состоящий из 76 аминокислотных остатков, обнаружен у многих организмов.