БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ - Е. С. Северина - 2004
РАЗДЕЛ 15. БИОХИМИЯ МЕЖКЛЕТОЧНОГО МАТРИКСА
У многоклеточных организмов большинство клеток окружено вне- или межклеточным матриксом. Межклеточный матрикс — сложный комплекс связанных между собой макромолекул. Эти макромолекулы (белки и гетерополисахариды), как правило, секретируются самими клетками, а в межклеточном матриксе из них строится упорядоченная сеть. Межклеточный матрикс, окружающий клетки, влияет на их прикрепление, развитие, пролиферацию, организацию и метаболизм.
Межклеточный матрикс вместе с клетками разного типа, которые в нём находятся (фибробласты, хондро- и остеобласты, тучные клетки и макрофаги), часто называют соединительной тканью.
Межклеточный матрикс выполняет в организме самые разнообразные функции:
✵ образует каркас органов и тканей;
✵ является универсальным «биологическим» клеем;
✵ участвует в регуляции водно-солевого обмена;
✵ образует высокоспециализированные структуры (кости, зубы, хрящи, сухожилия, базальные мембраны).
Основные компоненты межклеточного матрикса — структурные белки коллаген и эластин, гликозаминогликаны, протеогликаны, а также неколлагеновые структурные белки (фибронектин, ламинин, тенасцин, остеонектин и др.).
I. Коллаген
Коллаген — основной структурный белок межклеточного матрикса. Он составляет от 25 до 33% общего количества белка в организме, т. е. ~6% массы тела. Название «коллаген» объединяет семейство близкородственных фибриллярных белков, которые являются основным белковым элементом кожи, костей, сухожилий, хряща, кровеносных сосудов, зубов. В разных тканях преобладают разные типы коллагена, а это, в свою очередь, определяется той ролью, которую коллаген играет в конкретном органе или ткани. Например, в пластинчатой костной ткани, из которой построено большинство плоских и трубчатых костей скелета, коллагеновые волокна имеют строго ориентированное направление: продольное — в центральной части пластинок, поперечное и под углом — в периферической. Это способствует тому, что даже при расслоении пластинок фибриллы одной пластинки могут продолжаться в соседние, создавая таким образом единую волокнистую структуру кости. Поперечно ориентированные коллагеновые волокна могут вплетаться в промежуточные слои между костными пластинками, благодаря чему достигается прочность костной ткани. В сухожилиях коллаген образует плотные параллельные волокна, которые дают возможность этим структурам выдерживать большие механические нагрузки. В хрящевом матриксе коллаген образует фибриллярную сеть, которая придаёт хрящу прочность, а в роговице глаза коллаген участвует в образовании гексагональных решёток десцеметовых мембран, что обеспечивает прозрачность роговицы, а также участие этих структур в преломлении световых лучей. В дерме фибриллы коллагена ориентированы таким образом, что формируют сеть, особенно хорошо развитую в участках кожи, которые испытывают сильное давление (кожа подошв, локтей, ладоней), а в заживающей ране они агрегированы весьма хаотично. Аминокислотный состав и конформация коллагена описаны в подразделе «Фибриллярные белки» раздела 1.
Здесь будут разобраны синтез и созревание коллагена, структуры, которые он образует, и их функции, а также заболевания, связанные с нарушением этих процессов.
Полиморфизм коллагена
Коллаген — ярко выраженный полиморфный белок. В настоящее время известно 19 типов коллагена, которые отличаются друг от друга по первичной структуре пептидных цепей, функциям и локализации в организме. Вариантов α-цепей, образующих тройную спираль, гораздо больше 19 (около 30). Для обозначения каждого вида коллагена пользуются определённой формулой, в которой тип коллагена записывается римской цифрой в скобках, а для обозначения α-цепей используют арабские цифры: например, коллагены II и III типа образованы идентичными α-цепями, их формулы, соответственно [α1(II)]3 и [α1 (III)]3; коллагены I и IV типов являются гетеротримерами и образуются обычно двумя разными типами α-цепей, их формулы, соответственно [α1(I)]2α2(I) и [α1(IV)]2α2(IV). Индекс за скобкой обозначает количество идентичных α-цепей. Распределение коллагенов по органам и тканям представлено в табл. 15-1.
Таблица 15-1. Распределение коллагена в тканях и органах
Типы |
Гены |
Ткани и органы |
I |
СОL1А1, СОL1А2 |
Кожа, сухожилия, кости, роговица, плацента, артерии, печень, дентин |
II |
СОL2А1 |
Хрящи, межпозвоночные диски, стекловидное тело, роговица |
III |
СОL3А1 |
Артерии, матка, кожа плода, строма паренхиматозных органов |
IV |
СОL4А1-СОL4А6 |
Базальные мембраны |
V |
СОL5А1-СОL5А3 |
Минорный компонент тканей, содержащих коллаген I и II типов (кожа, роговица, кости, хрящи, межпозвоночные диски, плацента) |
VI |
СОL6А1-СОL6А3 |
Хрящи, кровеносные сосуды, связки, кожа, матка, лёгкие, почки |
VII |
СOL7А1 |
Амнион, кожа, пищевод, роговица, хорион |
VIII |
СОL8A1-СОL8А2 |
Роговица, кровеносные сосуды, культуральная среда эндотелия |
IX |
СOL9А1-СOL9А3 |
Ткани, содержащие коллаген II типа (хрящи, межпозвоночные диски, стекловидное тело) |
X |
СОL10А1 |
Хрящи (гипертрофированные) |
XI |
СОL1А1-СОL11А2 |
Ткани, содержащие коллаген II типа (хрящи, межпозвоночные диски, стекловидное тело) |
XII |
СОL12А1 |
Ткани, содержащие коллаген I типа (кожа, кости, сухожилия и др.) |
XIII |
СОL13А1 |
Многие ткани |
XIV |
СОL14А1 |
Ткани, содержащие коллаген I типа (кожа, кости, сухожилия и др.) |
XV |
СОL15А1 |
Многие ткани |
XVI |
СОL16А1 |
Многие ткани |
XVII |
СОL17А1 |
Гемидесмосомы кожи |
XVIII |
СОL18А1 |
Многие ткани, например печень, почки |
XIX |
СОL9А1 |
Клетки рабдомиосаркомы |
Гены коллагена называются соответственно типам коллагена и записываются арабскими цифрами, например, COL 1 — ген коллагена I типа, COL 2 — ген коллагена II типа, COL7 — ген коллагена VII типа и т. д. К этому символу приписываются буква А (обозначает α-цепь) и арабская цифра (обозначает вид α-цепи). Например, COL1A1 и COL1А2 кодируют, соответственно, α1 и α2-цепи коллагена I типа.
А. Этапы синтеза и созревания коллагена
Синтез и созревание коллагена — сложный многоэтапный процесс, начинающийся в клетке, а завершающийся в межклеточном матриксе. Синтез и созревание коллагена включают в себя целый ряд посттрансляционных изменений (рис. 15-1):
✵ гидроксилирование пролина и лизина с образованием гидроксипролина (Нур) и гидроксилизина (Нуl);
✵ гликозилирование гидроксилизина;
✵ частичный протеолиз — отщепление «сигнального» пептида, а также N- и С-концевых пропептидов;
✵ образование тройной спирали.
Рис. 15-1. Синтез и созревание коллагена.
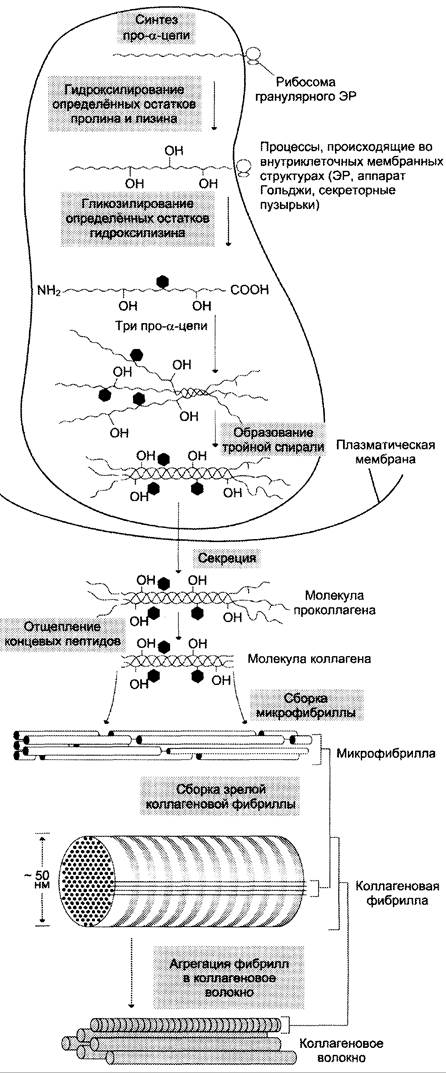
Синтез полипептидных цепей коллагена
Полипептидные цепи коллагена синтезируются на полирибосомах, связанных с мембранами ЭР, в виде более длинных, чем зрелые цепи, предшественников — препро-α-цепей. У этих предшественников имеется гидрофобный «сигнальный» пептид на N-конце, содержащий около 100 аминокислот.
Основная функция сигнального пептида — ориентация синтеза пептидных цепей в полость ЭР. После выполнения этой функции сигнальный пептид сразу же отщепляется. Синтезированная молекула проколлагена содержит дополнительные участки — N- и С-концевые пропептиды, имеющие около 100 и 250 аминокислот, соответственно. В состав пропептидов входят остатки цистеина, которые образуют внутри- и межцепочечные (только в С-пептидах) S-S-связи. Концевые пропептиды не образуют тройную спираль, а формируют глобулярные домены. Отсутствие N- и С-концевых пептидов в структуре проколлагена нарушает правильное формирование тройной спирали.
Посттрансляционные модификации коллагена
Гидроксилирование пролина и лизина. Роль витамина С
Гидроксилирование пролина и лизина начинается в период трансляции коллагеновой мРНК на рибосомах и продолжается на растущей полипептидной цепи вплоть до её отделения от рибосом. После образования тройной спирали дальнейшее гидроксилирование пролиловых и лизиловых остатков прекращается.
Реакции гидроксилирования катализируют оксигеназы, связанные с мембранами микросом. Пролиловые и лизиловые остатки в Y-положении пептида (Гли-х-у)n подвергаются действию, соответственно, пролил-4-гидроксилазы и лизил-5-гидроксилазы. Пролил-3-гидроксилаза действует на некоторые остатки пролина в Х-положениях. Необходимыми компонентами этой реакции являются α-кетоглутарат, O2 и витамин С (аскорбиновая кислота). Донором атома кислорода, который присоединяется к С-4 пролина, является молекула O2, второй атом O2 включается в сукцинат, который образуется при декарбоксилировании α-кетоглутарата, а из карбоксильной группы α-кетоглутарата образуется СO2(см. схему А).
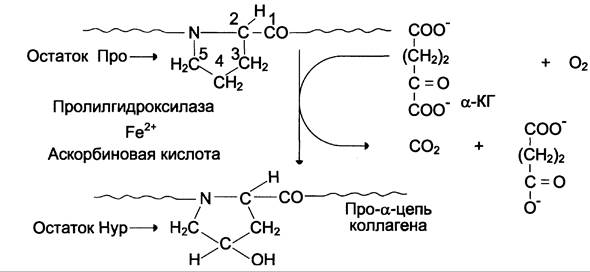
Гидроксилазы пролина и лизина содержат в активном центре атом железа Fе2+. Для сохранения атома железа в ферроформе необходим восстанавливающий агент. Роль этого агента выполняет кофермент гидроксил аз — аскорбиновая кислота, которая легко окисляется в дегидроаскорбиновую кислоту. Обратное превращение происходит в ферментативном процессе за счёт восстановленного глутатиона (см. схему Б).
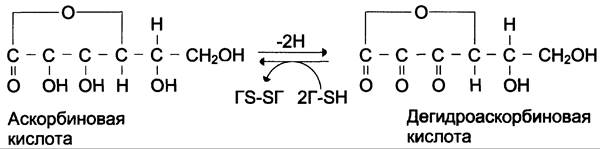
Гидроксилирование пролина необходимо для стабилизации тройной спирали коллагена, ОН-группы гидроксипролина (Нур) участвуют в образовании водородных связей. А гидроксилирование лизина очень важно для последующего образования ковалентных связей между молекулами коллагена при сборке коллагеновых фибрилл. При цинге — заболевании, вызванном недостатком витамина С, нарушается гидроксилирование остатков пролина и лизина. В результате этого образуются менее прочные и стабильные коллагеновые волокна, что приводит к большой хрупкости и ломкости кровеносных сосудов с развитием цинги. Клиническая картина цинги характеризуется возникновением множественных точечных кровоизлияний под кожу и слизистые оболочки, кровоточивостью дёсен, выпадением зубов, анемией.
Гликозилирование гидроксилизина
После завершения гидроксилирования при участии специфических гликозилтрансфераз в состав молекулы проколлагена вводятся углеводные группы. Чаще всего этими углеводами служат галактоза или дисахарид галакгозилглю- коза (рис. 15-2).
Рис. 15-2. Углеводные компоненты коллагена.
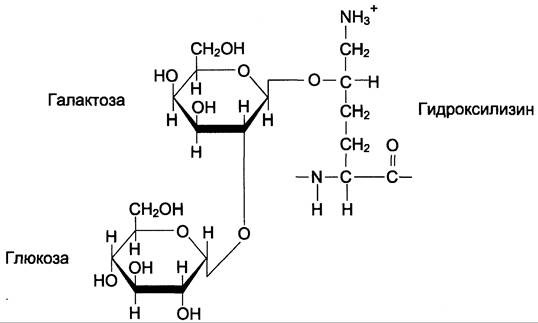
Они образуют ковалентную О-гликозидную связь с 5-ОН-группой гидроксилизина. Гликозилирование гидроксилизина происходит в коллагене, ещё не претерпевшем спирализации, и завершается после образования тройной спирали. Число углеводных единиц в молекуле коллагена зависит от вида ткани. Так, например, в коллагене сухожилий (тип I) это число равно 6, а в коллагене капсулы хрусталика (тип IV) — 110. Роль этих углеводных групп неясна; известно только, что при наследственном заболевании, причиной которого является дефицит лизилгидроксилазы (синдром Элерса — Данло — Русакова, тип VI), содержание гидроксилизина и углеводов в образующемся коллагене снижено; возможно, это является причиной ухудшения механических свойств кожи и связок у людей с этим заболеванием.
Образование проколлагена и его секреция в межклеточное пространство
После гидроксилирования и гликозилирования каждая про-α-цепь соединяется водородными связями с двумя другими про-α-цепями, образуя тройную спираль проколлагена. Эти процессы происходят ещё в просвете ЭР и начинаются после образования межцепочечных дисульфидных мостиков в области С-концевых пропептидов. Из ЭР молекулы проколлагена перемещаются в аппарат Гольджи, включаются в секреторные пузырьки и секретируются в межклеточное пространство.
Образование тропоколлагена. Болезни, связанные с нарушениями этого процесса
В межклеточном матриксе концевые пропептиды коллагенов I, II и III типов отщепляются специфическими проколлагенпептидазами, в результате чего образуются молекулы тропоколлагена, которые и являются структурной единицей коллагеновых фибрилл. При снижении активности этих ферментов (синдром Элерса-Данло-Русакова, тип VII) концевые пропептиды проколлагена не отщепляются, вследствие чего нарушается образование тропоколлагена и далее нарушается образование нормальных коллагеновых фибрилл. Нити коллагена видны под микроскопом в виде дезорганизованных пучков. Клинически это проявляется малым ростом, искривлением позвоночника, привычными вывихами суставов, высокой растяжимостью кожи.
У коллагенов некоторых типов (IV, VIII, X) концевые пропептиды не отщепляются. Это связано с тем, что такие коллагены образуют не фибриллы, а сетеподобные структуры, в формировании которых важную роль играют концевые N- и С-пептиды.
Б. Особенности структуры и функции разных типов коллагенов
19 типов коллагена подразделяют на несколько классов в зависимости от того, какие структуры они могут образовывать. Эти структуры представлены в табл. 15-2.
Таблица 15-2. Классификация коллагенов по видам структур, которые они образуют
Структура |
Тип |
Фибриллы |
I, II, III, V, XI |
Ассоциированные с фибриллами |
IX, XII, XIV, XVI, XIX |
Сети |
IV, VIII, X |
Микрофибриллы |
VI |
«Заякоренные» фибриллы |
VII |
Трансмембранные домены |
XIII, XVII |
Другие |
XV, XVIII |
Фибриллообразующие (I, II, III, V и XI) типы
95% всего коллагена в организме человека составляют коллагены I, II и III типов, которые образуют очень прочные фибриллы. Значительное содержание именно этих типов коллагена объясняется тем, что они являются основными структурными компонентами органов и тканей, которые испытывают постоянную или перио-
дическую механическую нагрузку (кости, сухожилия, хрящи, межпозвоночные диски, кровеносные сосуды), а также участвуют в образовании стромы паренхиматозных органов. Поэтому коллагены I, II и III типов часто называют интерстициальными. К классу фибриллообразующих относят также минорные коллагены V и XI типов.
Структура фибрилл коллагена и их формирование
Основа структурной организации коллагеновых фибрилл — ступенчато расположенные параллельные ряды молекул тропоколлагена, которые сдвинуты на 1/4 относительно друг друга (рис. 15-3).
Рис. 15-3. Схема ступенчатого расположения молекул коллагена в коллагеновой фибрилле.
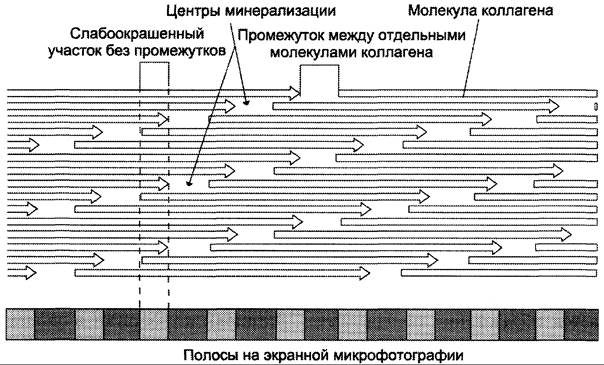
На схеме хорошо видно, что молекулы коллагена не связаны между собой «конец в конец», а между ними имеется промежуток в 35 — 40 нм. Предполагается, что в костной ткани эти промежутки выполняют роль центров минерализации, где откладываются кристаллы фосфата кальция. При электронной микроскопии фиксированные и контрастированные фибриллы коллагена выглядят поперечно исчерченными с периодом 67 нм, который включает одну тёмную и одну светлую полоски. Считают, что такое строение максимально повышает сопротивление всего агрегата растягивающим нагрузкам.
Фибриллы коллагена образуются спонтанно, путём самосборки. Но эти фибриллы ещё не являются зрелыми, так как не обладают достаточной прочностью (известно, что зрелое коллагеновое волокно толщиной в 1 мм выдерживает нагрузку до 10 кг).
Образовавшиеся коллагеновые фибриллы укрепляются внутри- и межцепочечными ковалентными сшивками (они встречаются только в коллагене и эластине). Эти сшивки образуются следующим образом:
✵ внеклеточный медьсодержащий фермент лизилоксидаза осуществляет окислительное дезаминирование ε-аминогрупп в некоторых остатках лизина и гидроксилизина с образованием реактивных альдегидов (аллизина и гидроксиаллизина). Для этих реакций необходимо присутствие витаминов РР и В6 (рис. 15-4).
Рис. 15-4. Образование поперечных связей в коллагене. А — образование альдольной поперечной сшивки из двух боковых цепей лизина; Б — образование шиффовых оснований из боковых цепей лизина и аллизина.
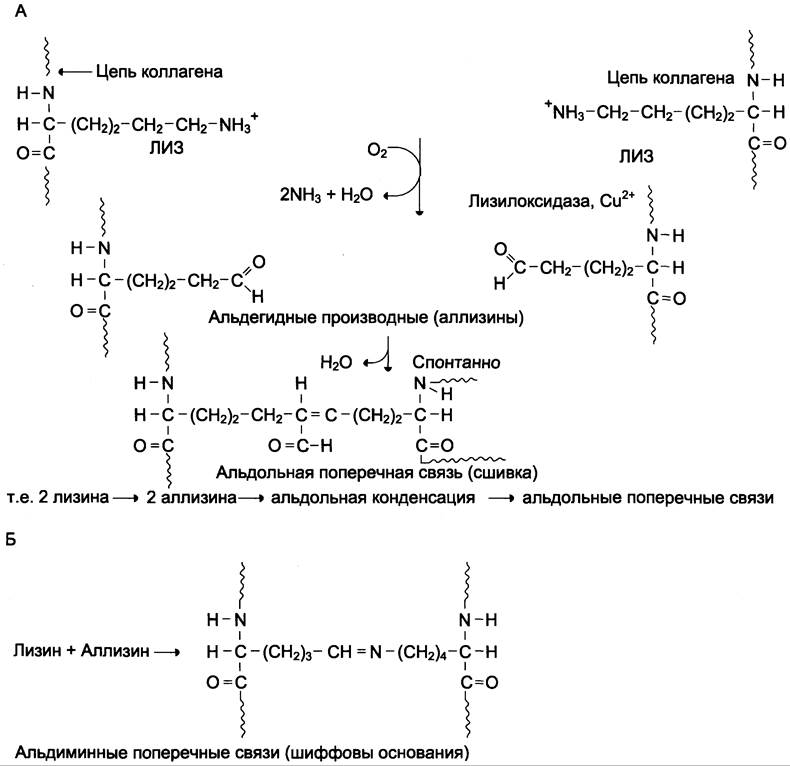
✵ образовавшиеся реактивные альдегиды участвуют в формировании ковалентных связей между собой, а также с другими остатками лизина или гидроксилизина соседних молекул тропоколлагена, и в результате возникают поперечные «Лиз-Лиз-сшивки», стабилизирующие фибриллы коллагена (рис. 15-5).
Рис. 15-5. Внутри- и межмолекулярные поперечные связи в коллагене.
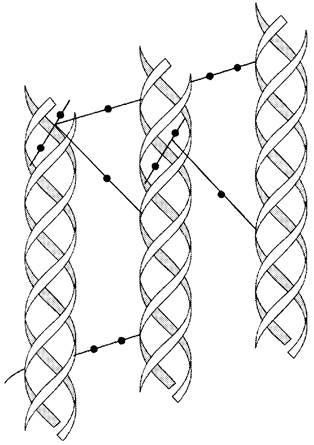
Количество поперечных связей в фибриллах коллагена зависит от функции и возраста ткани. Например, между молекулами коллагена ахиллова сухожилия сшивок особенно много, так как для этой структуры важна большая прочность. С возрастом количество поперечных связей в фибриллах коллагена возрастает, что приводит к замедлению скорости его обмена у пожилых и старых людей.
При снижении активности лизилоксидазы, а также при недостатке меди или витаминов РР или В6 нарушается образование поперечных сшивок и, как следствие, снижаются прочность и упругость коллагеновых волокон. Такие структуры, как кожа, сухожилия, кровеносные сосуды, становятся хрупкими, легко разрываются.
Подробнее эти вопросы рассматриваются ниже в подразделе, посвящённом эластину.
Коллагены, ассоциированные с фибриллами
Этот класс объединяет коллагены, которые выполняют очень важную функцию: они ограничивают размер фибрилл, образуемых интерстициальными коллагенами (прежде всего, I и II типов), и участвуют в организации межклеточного матрикса в костях, коже, хрящах, сухожилиях. К этим коллагенам относят коллагены IX, XII, XIV и XVI типов. Коллагены этого класса сами фибрилл не формируют, но непосредственно связаны с фибриллами, которые образуют интерстициальные коллагены. Функционирование таких типов коллагенов можно рассмотреть на примере коллагена IX типа, который в хряще связан с фибриллами коллагена II типа, он присоединяется к ним антипараллельно с периодичностью ~67 нм (рис. 15-6).
Рис. 15-6. Структура коллагеновых фибрилл II типа и ассоциированного с ним коллагена IX типа.
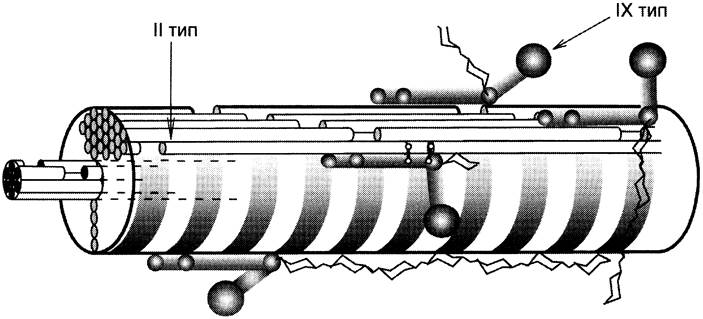
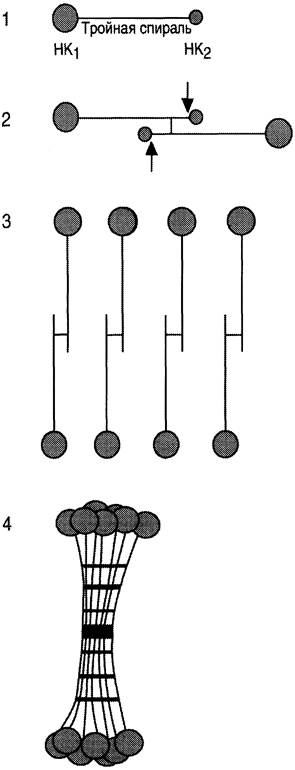
Коллаген IX типа состоит из трёх коллагеновых (фибриллярных) доменов (Кол1 —> Кол3) и четырёх неколлагеновых (глобулярных) доменов (НК1 —> НК4) (нумерация с С-конца) (рис. 15-7).
Рис. 15-7. Модель структуры коллагена IX типа. КОЛ1-КОЛ3 — коллагеновые домены; НK3-НК4 — неколлагеновые структуры.
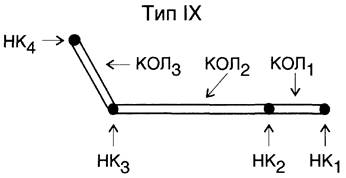
Коллаген IX типа связан с фибриллами коллагена II типа поперечными «Лиз-Лиз-мостиками» в области доменов Кол1 и Кол2, а также и НК1, НК2 и HK3.
НК4-домен не связан с фибриллами коллагена II типа; к его особенностям относят наличие большого количества положительно заряженных групп, поэтому к нему могут присоединяться отрицательно заряженные гликозаминогликаны, например, гиалуроновая кислота и хондроитин- сульфат. Эти особенности обеспечивают участие коллагена IX типа в организации межклеточного матрикса в хряще.
Коллагены, образующие сетеподобные структуры
К этому классу относят коллагены IV, VIII, X типов. Особенности строения и функционирования таких белков можно рассмотреть на примере наиболее изученных к настоящему времени коллагенов IV и VIII типов.
Коллаген IV типа является ключевым структурным компонентом базальных мембран, которые представляют собой особую форму межклеточного матрикса. Его секретируют различные типы клеток: эпителиальные, эндотелиальные, мышечные, нервные, жировые. Особенностью коллагена IV типа является то, что повторяющиеся спирализованные участки с последовательностью (Гли-х-у)n часто прерываются короткими неспиральными сегментами. Это, вероятно, увеличивает гибкость коллагена IV типа и способствует образованию на его основе сетчатых структур (рис. 15-8).
Рис. 15-8. Организация коллагена IV типа. А. Тройная спираль мономера коллагена: 7S — N-конец; НК1 — С-конец. Б. Полимеризация коллагена IV типа: 1 — мономер; 2 — димеры, образованные соединением мономеров в области НК1-доменов; 3 — тетрамеры, образованные соединением мономеров в области 7S-сегментов в параллельном и антипараллельном направлениях; 4 — образование сетчатой структуры из олигомерных форм коллагена IV типа.
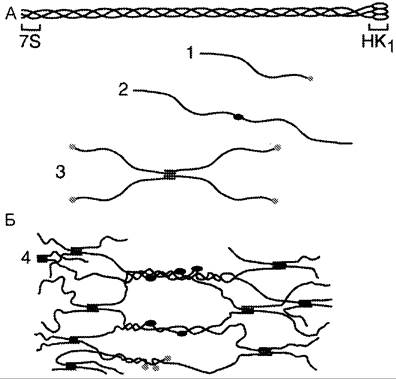
Молекулы этого коллагена не могут ассоциироваться латерально с образованием фибрилл, так как N- и С-концевые пропептиды у него не отщепляются. Но именно эти фрагменты участвуют в образовании олигомерных форм коллагена, так как они имеют ряд потенциальных мест связывания (остатки цистеина и лизина). Дисульфидные мостики и поперечные лизиновые связи стабилизируют образующиеся олигомеры. Кроме этого, возможны латеральные взаимодействия спирализованных участков разных молекул с образованием суперспиралей. В базальной мембране из этих компонентов формируется сетчатая структура с гексагональными ячейками размером 170 нм.
Коллагены VIII и X типов относят к так называемым короткоцепочечным коллагенам. Каждая их молекула состоит из короткого коллагенового домена, который составляет ~1/2 длины интерстициальных коллагенов, и неколлагеновых фрагментов на N- и С-концах.
Коллаген VIII типа — главный компонент десцеметовых мембран эндотелия роговицы. Молекулы этого коллагена собираются анти параллельно с образованием тетрамеров, из которых образуются гексагональные решётки, обеспечивающие прозрачность роговицы (рис. 15-9).
Рис. 15-9. Возможный механизм образования гексагональных решёток молекулами коллагена VIII типа. 1 — мономер; 2 — димер; 3 — тетрамер; 4 — гексагональные решётки.
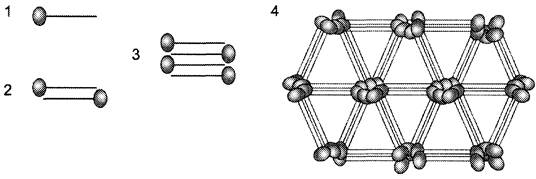
Кроме роговицы, коллаген VIII типа присутствует во многих других тканях, но ещё одна его преимущественная локализация — кровеносные сосуды, в которых он в основном находится в матриксе под эндотелиальными клетками. Образует ли этот коллаген и здесь гексагональные решётки, неизвестно. Возможно, что в сосудах коллаген VIII типа образует сетевидные структуры, подобные тем, которые формирует коллаген IV типа в базальных мембранах.
Коллагены, образующие микрофибриллы
К этому классу относят коллаген VI типа, который является короткоцепочечным белком. Он образует микрофибриллы, которые располагаются между крупными фибриллами интерстициальных коллагенов. Этот коллаген широко представлен в хрящевом матриксе, но больше всего его содержится в межпозвоночных дисках: в nucleus pulposus он составляет ~20% общего коллагена. Две молекулы этого коллагена соединяются антипараллельно с образованием димера. Из димеров образуются тетрамеры, которые секретируются из клетки, и вне клетки связываются «конец в конец» с образованием микрофибрилл (рис. 15-10).
Рис. 15-10. Организация коллагена VI типа. 1 — мономер; 2 — димер; 3 — тетрамер, соединённый полностью; 4 — тетрамер, соединённый частично; 5 — микрофибриллы, соединённые «конец в конец».
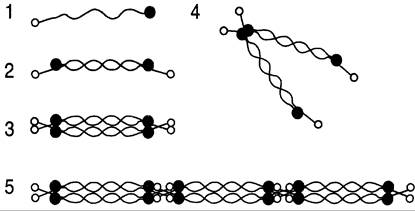
Функции коллагена VI типа пока полностью не ясны, хотя известно, что его микрофибриллы могут связываться со многими компонентами межклеточного матрикса: фибриллами интерстициальных коллагенов, гиалуроновой кислотой, протеогликанами. Молекула этого коллагена содержит многочисленные последовательности Арг-Гли-Асп (RGD), поэтому возможно его участие в клеточной адгезии через присоединение к мембранным адгезивным молекулам, например, интегринам α1β1 и α2β1.
Коллагены, образующие «заякоренные» фибриллы
К этому классу относят коллагены VII и XVII типов, которые называют также коллагенами, связанными с эпителием, так как они обычно находятся в местах соединения эпителия с субэпителиальными слоями.
Коллаген VII типа — основной структурный компонент «заякоренных» фибрилл. Каждая молекула этого белка содержит два неколлагеновых домена (НК1 — у С-конца, НК2 — у N-конца) и один коллагеновый домен между ними. Из мономеров образуются димеры, при этом молекулы соединяются в области НК2-доменов антипараллельно по отношению друг к другу. Затем НК2-домены отщепляются, и димеры соединяются между собой «бок о бок» с образованием фибрилл (рис. 15-11).
Рис. 15-11. Организация коллагена VII типа. 1 — мономер коллагена VII типа, НК1 и НК2 — неколлагеновые домены у С- и N-конца; 2 — димер коллагена VII типа, молекулы собраны антипараллельно с перекрытиями на N-конце; 3 — димеры коллагена VII типа после удаления НК2-доменов; 4 — фибрилла, образованная димерами коллагена VII типа, соединёнными «бок о бок».
Эти фибриллы играют важную роль в присоединении эпидермиса к дерме, так как одним концом они могут присоединяться к lamina densa, на которой лежит кожный эпителий, а другой их конец проникает в более глубокие субэпидермальные слои кожи и связывается там со структурами, называемыми «якорные диски».
Коллаген XVII типа представляет собой трансмембранный белок и обычно находится в гемидесмосомах эпидермиса. Предполагают, что этот коллаген взаимодействует с другими молекулами гемидесмосом и таким образом участвует в процессе присоединения эпидермиса к дерме.
В. Катаболизм коллагена
Как и любой белок, коллаген функционирует в организме определённое время. Его относят к медленно обменивающимся белкам; Т1/2 составляет недели или месяцы. Разрушение коллагеновых волокон осуществляется активными формами кислорода и/или ферментативно (гидролитически).
Коллагеназы, особенности их функционирования
Нативный коллаген не гидролизуется обычными пептидгидролазами. Основной фермент его катаболизма — коллагеназа, которая расщепляет пептидные связи в определённых участках спирализованных областей коллагена. Известны 2 типа коллагеназ.
Тканевая коллагеназа присутствует у человека в различных органах и тканях. В норме она синтезируется клетками соединительной ткани, прежде всего, фибробластами и макрофагами. Тканевая коллагеназа — металлозависимый фермент, который содержит Zn2+в активном центре. В настоящее время известно 4 изоформы этого фермента. Активность коллагеназы зависит от соотношения в межклеточном матриксе её активаторов и ингибиторов. Среди активаторов особую роль играют плазмин, калликреин и катепсин В (см. раздел 14). Тканевая коллагеназа обладает высокой специфичностью, она перерезает тройную спираль коллагена в определённом месте, примерно на 1/4 расстояния от С-конца, между остатками глицина и лейцина (или изолейцина) (см. схему ниже).
![]()
Образующиеся фрагменты коллагена растворимы в воде, при температуре тела они спонтанно денатурируются и становятся доступными для действия других протеолитических ферментов. Нарушение катаболизма коллагена ведёт к фиброзу органов и тканей (в основном печени и лёгких). А усиление распада коллагена происходит при аутоиммунных заболеваниях (ревматоидном артрите и системной красной волчанке) в результате избыточного синтеза коллагеназы при иммунном ответе.
Бактериальная коллагеназа синтезируется некоторыми микроорганизмами. Например, Clostridium histolyticum (возбудитель газовой гангрены) выделяет коллагеназу, расщепляющую пептидную цепь коллагена более чем в 200 местах. Этот фермент гидролизует следующую связь - X-Гли-Про-У- между звеньями X и Гли.
Таким образом разрушаются соединительнотканные барьеры в организме человека, что обеспечивает проникновение (или инвазию) этого микроорганизма и способствует возникновению и развитию газовой гангрены. Сам возбудитель не содержит коллагена и поэтому не подвержен действию коллагеназы.
Применение коллагеназ в медицине
Коллагеназа используется в медицинской практике для лечения ожоговой болезни в хирургии и для лечения гнойных заболеваний глаз в офтальмологии.
Определение гидроксипролина в физиологических жидкостях человека как показатель скорости распада коллагена
В результате распада коллагена в крови и моче появляется свободный гидроксипролин. Большая часть этой аминокислоты катаболизируется под действием фермента гидроксипролиноксидазы, а часть её выводится с мочой, и поэтому гидроксипролин является маркерной аминокислотой, по которой судят о скорости распада коллагена.
При некоторых заболеваниях, связанных с поражением соединительной ткани, экскреция гидроксипролина увеличивается вследствие ускоренного распада коллагена. Это наблюдается при болезни Педжета, гиперпаратиреозе, коллагенозах, некоторых инфекционных заболеваниях. При нарушении катаболизма гидроксипролина, причиной которого обычно выступает дефект фермента гидроксипролиноксидазы, выделение гидроксипролина может превышать 1 г/сут.
Особенности обмена коллагена
У молодых людей обмен коллагена протекает интенсивно, с возрастом (и особенно в старости) заметно снижается, так как у пожилых и старых людей увеличивается количество поперечных сшивок, что затрудняет доступность коллагена для действия коллагеназы. Поэтому, если у молодых людей в возрасте 10 — 20 лет содержание гидроксипролина в моче может достигать 200 мг/сут, то с возрастом экскреция гидроксипролина снижается до 15 — 20 мг/сут.
В некоторых ситуациях синтез коллагена заметно увеличивается. Например, фибробласты мигрируют в заживающую рану и начинают активно синтезировать в этой области основные компоненты межклеточного матрикса. Результат этих процессов — образование на месте раны соединительнотканного рубца, содержащего большое количество хаотично расположенных фибрилл коллагена. Сходным образом происходит замещение погибающих клеток соединительной тканью в печени при циррозе, в стенках артерий при атеросклерозе, в мышцах при их дистрофии.
Г. Регуляция обмена коллагена
Синтез коллагена регулируется разными способами. Прежде всего, сам коллаген и N-пропептиды после своего отщепления тормозят трансляцию коллагена по принципу отрицательной обратной связи. Аскорбиновая кислота стимулирует синтез коллагена и протеогликанов, а также пролиферацию фибробластов.
Особую роль в регуляции синтеза коллагена играют гормоны. Глюкокортикоиды тормозят синтез коллагена, во-первых, путём снижения уровня мРНК проколлагена, а во-вторых — ингибированием активности ферментов пролил- и лизилгидроксилазы. Недостаточное гидроксилирование остатков пролина и лизина повышает чувствительность коллагена к действию коллагеназы и неспецифических протеаз. Макроскопически угнетающее действие глюкокортикоидов на синтез коллагена проявляется уменьшением толщины дермы, а также атрофией кожи в местах продолжительного парентерального введения этих гормонов.
На синтез коллагена влияют также половые гормоны, рецепторы к которым обнаружены не только в строме половых органов, но и в фибробластах других органов и тканей. Обмен коллагена в матке находится под контролем половых гормонов. Синтез коллагена кожи зависит от содержания эстрогенов, что подтверждает тот факт, что у женщин в менопаузе снижается содержание коллагена в дерме.
Д. Заболевания, связанные с нарушением синтеза и созревания коллагена
Существует ряд заболеваний, связанных с нарушением структуры или синтеза коллагена. Основная причина — мутации в генах коллагена, которые широко представлены в разных хромосомах. Они очень большие, имеют много коротких экзонов, между которыми располагаются большие интроны.
Так как около 50% всех коллагеновых белков содержится в тканях скелета, около 40% — в коже и 10% — в строме внутренних органов,
клиническая картина заболеваний, вызванных дефектами синтеза и созревания коллагена, будет крайне полиморфной. При многих заболеваниях наблюдают не только костно-суставную патологию или изменения со стороны кожи, но и ярко выраженные висцеральные проявления (поражения кишечника, почек, лёгких, сердца, сосудов).
К настоящему времени описано много наследственных заболеваний, причинами которых являются дефекты коллагенов разных типов (см. ниже табл. 15-3).
Таблица 15-3. Заболевания, связанные с нарушением синтеза и созревания коллагена
Тип коллагена |
Ген |
Локализация коллагена в тканях |
Заболевания |
Причина |
Клинические проявления |
I |
СОLА1 СОL1А2 |
Кости, кожа, связки, сухожилия, склера, роговица, строма внутренних органов |
Несовершенный остеогенез |
Мутации в генах (более 160), чаще всего делеции и замены. Самая неблагоприятная — замена глицина на другую аминокислоту, в результате чего в молекуле проколлагена появляется перелом или изгиб, и нормальная тройная спираль не образуется |
Повышенная ломкость костей, аномалии зубов, треугольная форма лица, гиперподвижность суставов, голубые склеры |
II |
СОL2А1 |
Хрящи, межпозвоночные диски, стекловидное тело |
Болезнь Книста Синдром Стиклера и Вагнера |
Делеция в гене, которая приводит к синтезу укороченных цепей коллагена Образование терминирующего кодона, вследствие чего в стекловидном теле синтезируется половина молекулы коллагена |
Укорочение и деформации конечностей, тугоподвижность суставов, кифосколиоз, миопия высокой степени Прогрессирующая миопия, часто отслойка сетчатки; патология суставов по типу хронического остеоартрита |
III |
СОL3А1 |
Кожа, сосуды, строма паренхиматозных органов, матка |
Синдром Элерса- Данло- Русакова, IV тип |
Мутации в гене (более 20) по типу делеций, вставок, замен. В результате этого синтезируется молекула коллагена с нарушением первичной структуры, которая отличается сниженной стабильностью. Фибриллы, которые образуют такие молекулы коллагена, тоньше нормальных и менее организованы |
Спонтанные разрывы крупных сосудов, перфорации кишечника, разрывы беременной матки, спонтанный пневмоторакс |
IV |
СОL4А3- СОL4А6 |
Базальные мембраны (почки и лёгкие) |
Синдром Альпорта Синдром Гудпасчера |
Мутации в генах, которые сопровождаются нарушением образования базальных мембран Образование антител к молекулам коллагена IV типа |
Преимущественное поражение почек, проявляющееся гематурией и протеинурией; при некоторых формах одновременно развивается диффузный эзофагеальный лейомиоматоз (доброкачественная опухоль гладких мышц пищевода). Гломерулонефрит, лёгочный гемосидероз |
VII |
СOL7А1 |
Кожа |
Буллёзный эпидермолиз |
Мутации в гене, приводящие к снижению общего количества «заякоренных» фибрилл в коже, а также синтез дефектных фибрилл |
Эпидермис слабо связан с дермой, легко слущивается и образует пузыри (буллы), которые легко травмируются, и на их месте образуются эрозии |