Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ VI. БІОХІМІЯ ФІЗІОЛОГІЧНИХ ФУНКЦІЙ ТА СПЕЦІАЛІЗОВАНИХ ТКАНИН
ГЛАВА 27. БІОХІМІЯ ХАРЧУВАННЯ ЛЮДИНИ. II. ВІТАМІНИ ЯК КОМПОНЕНТИ ХАРЧУВАННЯ
27.3. ЖИРОРОЗЧИННІ ВІТАМІНИ. БІОАНТИОКСИДАНТИ
Вітаміни, що входять до цього класу, є олієподібними речовинами, які добре взаємодіють із гідрофобними розчинниками; завдяки наявності в структурі молекул довгих вуглеводневих (переважно ізопреноїдних) радикалів, більшість із цих вітамінів є компонентами біомембран, у складі яких виконують специфічні біологічні функції, зокрема є потужними біоантиоксидантами(вітаміни Е, А, К).
Всмоктування жиророзчинних вітамінів у кишечнику залежить від наявності поверхнево-активних компонентів жовчі і може порушуватися при обтурації жовчних проток, що супроводжується симптомами вітамінної недостатності. З іншого боку, на відміну від водорозчинних вітамінів, надлишкове (щодо фізіологічних потреб) надходження жиророзчинних вітамінів (особливо А, Д, К) є небезпечним для організму людини, оскільки ці сполуки можуть накопичуватися в тканинних депо і спричиняти токсичну дію (стан гіпервітамінозу).
Вітамін А
Хімічна будова та біологічні джерела
Сполуки, що мають біологічні властивості вітаміну А, є похідними β-іонону із загальною формулою:
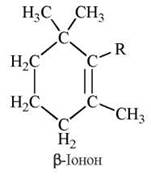
Дві молекулярні форми вітаміну А (вітамери) — А1 та А2 є циклічними ненасиченими спиртами (трансізомери), що мають як бічний радикал гідрофобну діізопреноїдну групу, завдяки якій ці сполуки розчиняються в ліпідному бішарі мембран:
Обидві сполуки проявляють повний спектр біологічних ефектів вітаміну А, проте вітамін А1 є дещо активнішим.
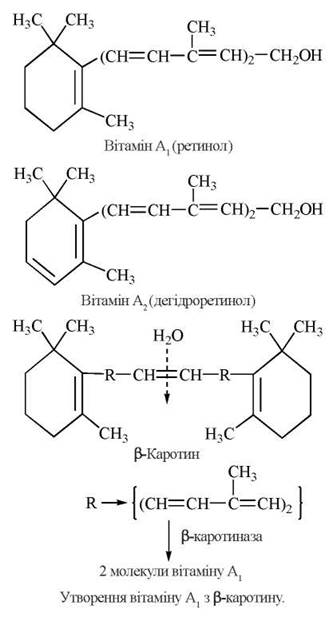
У рослинних організмах містяться провітаміни (біологічні попередники) вітаміну А — жовті пігменти α, β та γ-каротини (вперше були виявлені в моркві — carota; лат.). Найбільш активним провітаміном вітаміну А є β-каротин, при гідролізі якого за участю ферменту β-каротинази стінки тонкої кишки та печінки людини утворюються дві молекули вітаміну А1:
Біологічні властивості
Після надходження в організм людини (з тваринною їжею або у вигляді рослинних каротинів) ретинол та дегідроретинол депонуються в тканинах (переважно в печінці) у вигляді складних жирнокислотних ефірів, які, у міру фізіологічної потреби, утворюють активні молекулярні форми вітаміну А: спирт (ретинол), альдегід (ретиналь) таретиноєву кислоту:
![]()
Біологічна активність вітаміну А полягає, переважно, в регуляції таких функцій організму:
- процесів темнового (нічного) зору — недостатність вітаміну А супроводжується порушенням темнового зору і розвитком «курячої сліпоти» (гемералопії);
- процесів росту та диференціювання клітин;
- процесів утворення глікопротеїнів, що є компонентами біологічних слизів організму.
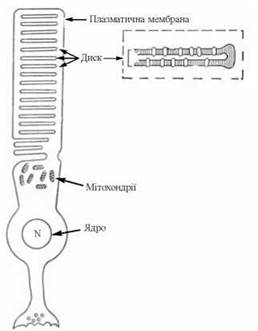
Рис. 27.3. Схема структурної організації дисків зовнішніх сегментів паличок сітківки.
1. Процеси темнового зору є фізіологічною функцією спеціалізованих клітин сітківки — паличок, яка забезпечується фоторецепторним білком родопсином, що міститься в мембранних утвореннях зовнішніх сегментів паличок — дисках (вп’ячуваннях плазматичної мембрани клітини).
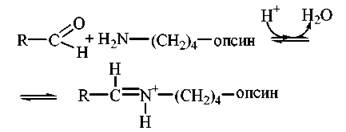
Родопсин — складний білок (м.м. 40 кД), що складається з білкової частини — опсину та хромофору (простетичної групи) — альдегідної форми вітаміну А — 11-цис-ретиналю, яка зв’язана з ε-аміногрупою лізинового залишку альдимінним зв’язком (основа Шиффа):
Будучи інтегральним (трансмембранним) білком мембрани диска, завдяки циклічним внутрішньомолекулярним перетворенням вітаміну А родопсин сприймає квант (один фотон!) світла, трансформуючи його в гіперполяризацію мембрани, тобто запускає елементарний фізіологічний акт зору. Послідовність молекулярних подій у цьому процесі є такою:
(1) сприйняття родопсином кванта світла (максимум поглинання при 500 нм) ініціює ізомеризацію зв’язаного з опсином 11-цис-ретиналю на повністю трансретиналь;
(2) фотоіндукована ізомеризація простетичної групи (вітаміну А) спричиняє зміни конформації білкової частини молекули і утворення декількох проміжних інтермедіатів (конформерів) родопсину: батородопсин — люміродопсин — метародопсин I — метародопсин II;
(3) останній з індукованих світлом конформерів родопсину — метародопсин II (т.з. «фотозбуджений родопсин» — R*) індукує каскад біохімічних реакцій, що призводять до закриття Na+ — каналів мембран дисків. Цей процес є каскадним механізмом передачі хімічного сигналу від збудженого родопсину через білок трансдуцин (аналог G-білка) на фосфодіестеразу, що гідролізує цГМФ; зменшення рівня цГМФ призводить до закриття Na+-каналу і гіперполяризації мембрани; гіперполяризація, що настає, є сигналом для подальшого електрохімічного реагування нейронів сітківки;
(4) транс-ретиналь, що утворився в результаті дії світла (п.1) втрачає зв’язок з білковою частиною родопсину і підлягає (в темряві) регенерації до 11-цис-ретиналю, який знову вступає в сполучення з лізиновим залишком білка, утворюючи функціонально активний родопсин (рис. 27.4).
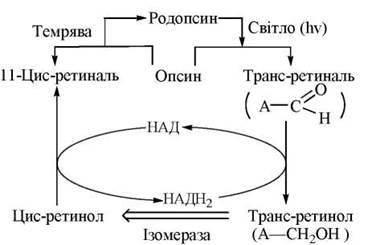
Рис. 27.4. Схема фотоіндукованих перетворень ізомерних форм вітаміну А в паличках сітківки.
2. З самого початку вивчення вітаміну А була встановлена його унікальна стимулювальна дія відносно процесів росту та диференціювання клітин («вітамін росту»). Згідно із сучасними уявленнями, ця біологічна функція реалізується транс-ретиноєвою кислотою (РК), що утворюється в організмі з альдегідної форми вітаміну А.
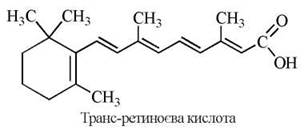
В основі стимулювання вітаміном А процесів росту та розвитку організмів (морфогенезу) лежить вплив транс-ретиноєвої кислоти на процеси транскрипції, молекулярні механізми яких були розглянуті в главі 23. Як випливає із зазначеного матеріалу, ядерні рецептори для РК належать до суперсімейства регуляторів транскрипції разом з рецепторами для стероїдних гормонів, вітаміну D3 та тироксину, молекулярно-генетичні механізми функціонування яких є об’єктами сучасних досліджень.
3. Характерним проявом недостатності вітаміну А у людини та тварин є виражена сухість слизових оболонок, вкритих одношаровим плоским епітелієм, що вистилає шлунково-кишковий та дихальний тракт, сечовивідні та статеві шляхи, очне яблуко, сльозний та слуховий канали тощо. Введення препаратів вітаміну А або продуктів, що його містять, протидіє вказаним патологічним проявам, зокрема сухості очного яблука («антиксерофтальмічний» вітамін, «аксерофтол»).
Біохімічною основою цієї групи ефектів вітаміну А є його стимулювальна дія відносно біосинтезу глікопротеїнів, які складають основу муцинів — слизових утворень, які вкривають зазначені епітеліальні покриви. Існують дані щодо участі вітаміну А у функціонуванні глікозилтрансфераз ендоплазматичного ретикулума та комплексу Гольджі, а саме коферментної функції ретинолу як ліпідного переносника олігосахаридних залишків через ліпопротеїнові мембрани до місць глікозилювання пептидної частини глікопротеїну.
Добова потреба
Вітамін А надходить в організм людини з продуктами тваринного походження (особливо у складі вершкового масла, сметани, молока, печінки, риб’ячого жиру, яєчного жовтка) та у вигляді рослинних каротинів.
Добова потреба у вітаміні А складає 1,5-2,5 мг, або 3-5 мг каротинів* [1 мг ретинолу, або 6 мг каротинів]**.
Вітамін К
Хімічна будова
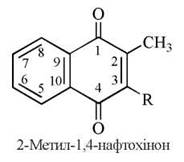
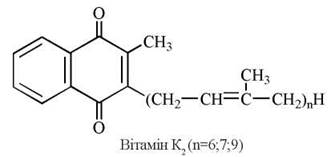
Властивості вітаміну К має група вітамерів — похідних 2-метил-1,4-нафтохінона. Розрізняють вітамін К1 (філохінон) та вітамін К2 (фарнохінон).
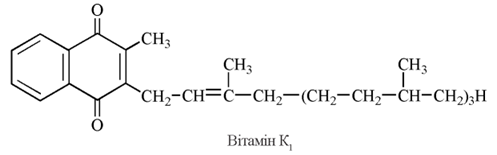
Вітамін К2 є похідним 2-метил-1,4-нафтохінону, бічним вуглеводневим радикалом, у якому є похідне ізопрену — фітил (2-метил-3-фітил-1,4-нафтохінон). Цей вітамер був вперше виділений із люцерни і є біологічно найбільш активною формою вітаміну К.
Вітамін К2 має довший бічний ізопреноїдний ланцюг, будучи за хімічною будовою 2-метил-3-фарнезил-1,4-нафтохіноном; вітамер був вперше виділений із рибного борошна.
Біологічні властивості
Біологічна дія вітаміну К в організмі людини і тварин полягає в його впливі на функціонування згортальної системи крові («антигеморагічний» вітамін). Оскільки вітамін К є необхідним компонентом для утворення факторів коагуляції крові II, VII, IX, X, недостатність вітаміну супроводжується небезпечними для життя кровотечами. Біохімічні механізми прокоагулянтної дії вітаміну К розглянуті в главі 29.
Гіповітаміноз вітаміну К у людини розвивається найчастіше при захворюваннях печінки та системи жовчовивідних шляхів, які перешкоджають утворенню та/або надходженню в дванадцятипалу кишку жовчі, необхідної для всмоктування жиророзчинних речовин. При підвищеній активності згортальної системи крові нагальною проблемою клінічної практики є застосування антикоагулянтів, що за механізмом дії є антивітамінами вітаміну К (група похідних кумарину).
Джерела та добова потреба
Джерелами вітаміну К для організму людини є переважно рослинні продукти харчування (капуста, помідори, салат); певна кількість вітаміну міститься в печінці (особливо свиній), м’ясі. Значна кількість вітаміну синтезується також кишковою мікрофлорою, що може забезпечити потреби організму людини в цьому вітаміні навіть в умовах зменшеного його надходження з продуктами харчування.
Добова потреба у вітаміні К складає 200-300 мкг* [80 мкг]**.
Вітамін Е
Хімічна будова
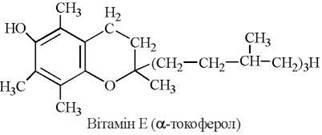
Властивості вітаміну Е має група похідних токолу (2-метил-2(4',8',12'-триметилтридецил)-6-хроманолу — α, β та γ-токофероли, що були вперше виділені з рослинних олій. Найбільшу біологічну активність має α-токоферол (5,7,8-триметилтокол):
Біологічні властивості та механізм дії
Вітамін Е має широкий спектр біологічної активності — його недостатність супроводжується численними змінами обмінних процесів та фізіологічних функцій організму. Найбільш характерними для Е-авітамінозу є глибокі порушення репродуктивної функції як у чоловіків (аномальний сперматогенез) так і жінок (неспроможність запліднення та виношування вагітності), м’язові дистрофії, некрозо-дистрофічні процеси в печінці.
Згідно з сучасними уявленнями, основні молекулярні механізми дії вітаміну Е (α-токоферолу) полягають у наступному:
(1) завдяки наявності вільного фенольного гідроксилу в ароматичному ядрі хроману α-токоферол може вступати в реакцію диспропорціонування з вільними радикалами у вигляді гасника (інгібітора) вільних радикалів InH, гальмуючи процеси вільно-радикального окислення органічних молекул:
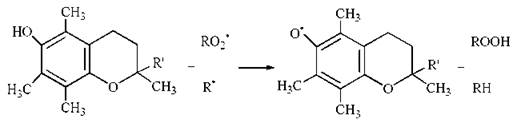
Продукти реакції α-токоферолу з органічними радикалами (In’) вступають в реакцію між собою, утворюючі неактивні молекулярні продукти In-In:
![]()
(2) завдяки гідрофобному бічному радикалові α-токоферол може вбудовуватися в фосфоліпідний матрикс біомембран, стабілізуючи рухомість та мікров’язкість мембранних ліпідів і білків.
Антирадикальні та мембраностабілізуючі властивості вітаміну Е є біохімічною основою його біологічної функції як найбільш потужного біоантиоксиданта. Протидіючи перекисному окисленню біомолекул (ліпідів, білків, нуклеїнових кислот), α-токоферол захищає клітинні структури від цитотоксичної дії вільних радикалів як ендогенного походження, так і ксенобіотиків, що потрапляють в організм із зовнішнього середовища (Ю.І. Губський, 1995).
Джерела та добова потреба
Найбільш багатими джерелами вітаміну Е в харчуванні людини є олії (соняшникова, кукурудзяна, соєва тощо), свіжі овочі та тваринні продукти (м’ясо, вершкове масло, яєчний жовток).
Добова потреба у вітаміні Е (α-токоферолі) становить 10-20 мг* [10 мг]**.
Вітамін F
Хімічна природа та властивості
Під вітамінами групи F розуміють групу поліненасичених жирних кислот рослинного походження — переважно лінолевої та ліноленової, що є попередниками у синтезі біологічно активних ейкозаноїдів — похідних арахідонової кислоти (простагландинів, тромбоксанів, лейкотрієнів).
Джерела та добова потреба
Джерелами поліненасичених жирних кислот є здебільшого рослинні олії, в деякій мірі — тваринні жири, вершкове масло, яйця. Добова потреба організму людини у вітаміні F складає близько 2-6 г.
Вітамін D
Хімічна будова
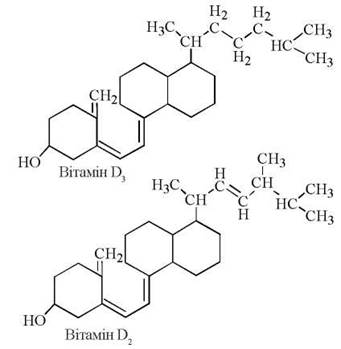
До вітамінів групи D належать вітамери D3 (холекальциферол, вітамін D тваринного походження) та D2 (ергокальциферол, вітамін D рослинного походження):
Біологічні властивості та механізм дії
Біологічною функцією вітамінів групи D є регуляція гомеостазу кальцію. Холекальциферол — вітамін D3, що утворюється в організмі людини з 7-дегідрохолестерину, є попередником фактора гормонального типу дії кальцитріолу (1,25(OH)2D3), який індукує синтез Са-зв’язуючих білків ентероцитів і є, таким чином, основним регулятором всмоктування в кишечнику іонів Са2+, необхідних для кісткоутворення та контролю багатогранних Са-залежних біохімічних процесів (глава 25).
Найбільш частими причинами недостатності вітаміну Д з порушенням кальцієво-фосфорного обміну, остеомаляцією і розвитком рахіту (rhachis — хребет; спинномозковий стовбур — грецьк.) у дітей є знижене сонячне опромінення шкіри, а також зменшене споживання тваринних продуктів, що містять холекальциферол.
Джерела та добова потреба
Найбільша кількість вітаміну D (D3) міститься в продуктах харчування тваринного походження: вершковому маслі, жовтку яєць, печінці; особливо багатим джерелом вітаміну D3 є риб ’ячий жир, що широко використовується для профілактики і лікування рахіту.
Антирахітну активність має також ергокальциферол (вітамін D2) що утворюється при ультрафіолетовому опроміненні рослинного стерину — ергостерину, який міститься в значній кількості в дріжджах та грибах.
Добова потреба в вітаміні D для дорослої людини складає 2,5-10 мкг* [5 мкг]**. Для дітей раннього віку — в середньому 12-25 мкг (Т.Т. Березов, Б.Ф. Коровкин, 1983); за рекомендаціями Ради з харчових продуктів та харчування Національної академії наук США — 7,5-10 мкг.