Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ IV. МОЛЕКУЛЯРНІ МЕХАНІЗМИ СПАДКОВОСТІ ТА РЕАЛІЗАЦІЇ ГЕНЕТИЧНОЇ ІНФОРМАЦІЇ
ГЛАВА 20. МОЛЕКУЛЯРНІ МЕХАНІЗМИ РЕПЛІКАЦІЇ ДНК ТА ТРАНСКРИПЦІЇ РНК
20.1. БІОЛОГІЧНЕ ЗНАЧЕННЯ РЕПЛІКАЦІЇ ДНК. НАПІВКОНСЕРВАТИВНИЙ МЕХАНІЗМ РЕПЛІКАЦІЇ
Значення відкриття в 1953 р. Дж. Уотсоном та Ф. Кріком будови молекули ДНК як подвійної спіралі полягало не тільки в розшифровці геометричної структури молекули. Модель, що була запропонована, розкривала можливості для створення чітких наукових уявлень про механізм подвоєння — реплікації молекули ДНК, тобто рівного розподілу між дочірніми клітинами генетичного матеріалу, що становило чи не найбільшу біологічну таємницю протягом усього існування людства.

Рис. 20.1. Уотсон (Watson) Джеймс Д. (народ. 1928 р.), американський біохімік, один із фундаторів молекулярної біології. Нобелівська премія (1962).

Рис. 20.2. Крік (Crick) Френсис (народ. 1916 р.), англійський фізик, один із фундаторів молекулярної біології. Нобелівська премія (1962).
Як зазначав пізніше Дж. Уотсон, «існування двох переплетених ланцюгів з однаковою послідовністю основ не могло бути випадковим. Навпаки, це дає право вважати, що один із ланцюгів кожної молекули на певній стадії править за матрицю для синтезу іншого ланцюга. За такою схемою реплікація гена починалася б із його розділення на два однакові ланцюги. Потім на обох материнських матрицях могли б утворюватися два дочірніх ланцюги, в результаті чого формувалися б дві молекули ДНК, ідентичні первісній» (J. Watson, 1968).
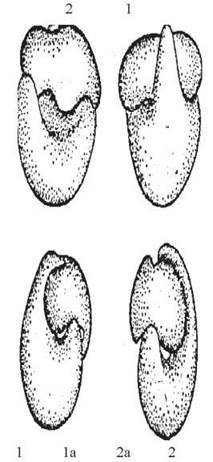
Рис. 20.3. Модель реплікації ДНК за механізмом Уотсона та Кріка.
1, 2 — материнські ланцюги ДНК; 1а, 2а — дочірні ланцюги ДНК.
Напівконсервативний та консервативний механізми реплікації
Модель подвоєння (реплікації) молекули ДНК, що була первинно запропонована Дж.Уотсоном та Ф .Кріком, отримала назву напівконсервативного механізму реплікації. Згідно з цим механізмом, «материнська» молекула ДНК в процесі реплікації розділяється на два ланцюги, кожен з яких править за матрицю для синтезу за принципом комплементарності другого ланцюга з утворенням ідентичних «дочірніх» молекул.
Поряд із зазначеною моделлю Уотсона-Кріка, як теоретично можливий розглядався також консервативний механізм реплікації, відповідно до якого при подвоєнні клітини «стара» материнська молекула ДНК залишається незайманою, а з вільних мононуклеотидів складаються повністю нові молекули ДНК:
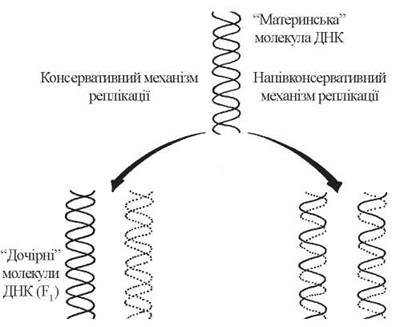
Рис. 20.4. Теоретично можливі механізми реплікації ДНК.
Експериментальне обґрунтування напівконсервативного механізму реплікації ДНК було здійснено в дослідах М. Мезелсона та Ф. Сталя (M. Meselson, F.W. Stahl; 1957). Головна ідея експерименту М. Мезелсона та Ф. Сталя ґрунтувалася на різній питомій щільності і, відповідно, можливості фракціонування методом ультрацентрифугування в градієнті щільності CsCl молекул ДНК, що відрізняються за вмістом легкого (N14) або важкого (N15) ізотопів азоту. Схему експерименту можна розділити на такі етапи:
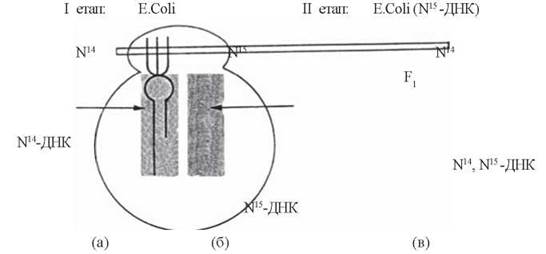
Рис. 20.5. Схема експерименту М. Мезелсона та Ф. Сталя.
1) вирощування клітин E. Coli на середовищі, що містило як джерело азоту NH4Cl з легким ізотопом (N14); була встановлена щільність «легких» молекул ДНК, що містилися в таких клітинах (рис. 20.5 а);
2) вирощування клітин E. Coli протягом декількох поколінь на середовищі, що містило як джерело азоту NH4Cl з важким ізотопом (N15); була встановлена щільність «важких» молекул ДНК, що містилися в таких клітинах (рис. 20.5б);
3) клітини E. Coli, що містили у своєму складі «повністю важкі» молекули ДНК (N15-ДНК) знову були перенесені на середовище з легким (N14) ізотопом азоту; після першої реплікації дочірні клітини (F1) збиралися, з них екстрагувалася ДНК і методом ультрацентрифугування визначалася її щільність. Було встановлено, що щільність цієї ДНК з покоління F1 була проміжною між щільністю «легких» (N14) та «важких» (N15) молекул ДНК, тобто складалася наполовину з «легких» і наполовину з «важких» ланцюгів, що відповідало будові (N14, N15-ДНК) (рис. 20.5в), підтверджуючи тим самим напівконсервативний механізм реплікації.